
文字/编辑:一口半夏
«——【 ·前言· 】——»
三种氨基酸环延伸同源域转录因子(TALE HD TFs)在绿藻和陆生植物中扮演着生存周期调节者的角色,在苔藓发育过程中,这些调控因子在孢子体的发育程序中发挥着作用。

本研究结果强调了在褐藻Ectocarpus中两个TALE HD TF基因的突变,(即OUROBOROS或SAMSARA)导致孢子体世代转化为配子体,这两种基因的异二聚体形式与绿色藻类中的TALE HD TF生命周期调控因子的作用方式相似。

本实验的最终目的在于揭示:TALE-HD-TF为基础的生命周期调节系统具有古老的起源,这些系统在至少两个不同的复杂多细胞真核超群体(古质体和Chromalveolata)中独立进化,并被招募来调控孢子体的发育计划,以调控孢子体的发育过程。

 两个同源结构域转录因子直接影响
两个同源结构域转录因子直接影响奥罗基因在本研究中被定位于14号染色体的一个34.5 kbp(0.45厘米)间隔上,利用了一个由2000个兄弟姐妹构成的分离家庭,该家庭源自奥罗x奥罗的杂交,并利用扩增片段长度多态性(AFLP)和微卫星标记进行了分析。

对位于中间34.5 kbp间隔的奥罗基因突变体进行的重测序表明,该突变体仅包含一个11 bp的缺失,位于基因Ec-14_005920的外显子6中,该基因编码一个TALE同源结构域的转录因子。
通过对约14,000个紫外线诱变细菌的视觉筛选,进一步鉴定了另外三个生命周期突变体(指定)。

其中山姆突变体在单性卵子中表现出与奥罗类似的特征,与正常孢子体发育模式不同,呈现出典型的配子体的花丝生长模式。
成熟时,山姆突变体不产生单眼孢子囊(减数分裂发生的生殖结构),这在孢子体世代期间是唯一观察到的结构。此外,山姆突变体对单侧光的负光营养反应较野生型更为明显,这是配子体的典型特征,奥罗突变体也表现出类似的现象。

外果皮孢子体会产生一种扩散因子,诱导配子体初始细胞或成熟配子体细胞向孢子体发育路径转变,巨兽突变体对这种扩散因子的敏感性较低(奥罗原生质体在孢子体培养基中表现为配子体再生),表明ORO在扩散因子的作用中起到了关键作用,直接参与孢子体发育途径的调控。

萨姆-1突变体对扩散因子的反应也表现出抗性,萨姆-1的原生质体在扩散因子处理下未检测到孢子体形成,而野生型配子体原生质体的对照处理导致7.5%的个体转化为孢子体,因此,对于扩散因子的响应需要细胞具备oro和萨姆。

Ectocarpus基因组中包含两个TALE HD TFs,除了奥罗基因,对这些基因的重测序在三个山姆突变体中鉴定出了3个基因突变,这些突变被预测会严重影响基因Ec-27_006660的功能。
同一基因中的三个破坏性突变在三个独立的山姆突变体中的鉴定强烈表明,这些突变是致病性的。因此,基因Ec-27_006660被命名为"轮回"。

ORO和SAM在配子中表达量最为丰富,与配子融合后孢子体发育的启动机制保持一致。
值得注意的是,转录本分析揭示,在雄配子和雌配子中均检测到ORO和SAM的存在,这意味着男性和女性配子都携带有Oro和SAM蛋白。
与此相比,与衣藻中的情形似乎不同,衣藻中GSP1和GSM1仅在正负配子中表达。

尽管我们不能排除转录后调控可能对ORO表达的影响,但山姆的性别特异性配子表达模式以及遗传学证据支持了双性表达的情况,至少对于ORO来说,这是因为携带oro突变的雄性和雌性菌株与野生型菌株表现出相似的性状。
通过定量PCR实验,我们进一步证实在sam突变系中,孢子体和配子体标记基因的表达分别被下调和上调,这与之前针对oro突变体的研究结果相一致。

 奥罗 还有 山姆 调控孢子体发生基因
奥罗 还有 山姆 调控孢子体发生基因为了深入探究基因调控对配子体向孢子体转化遗传机制的影响,我们系统性地描述了与 Ectocarpus 生命周期两个关键阶段相关的基因表达网络,即奥罗和萨姆基因。
通过对在相同生长条件下培养的野生型孢子体和配子体的重复培养物进行 RNA-seq 数据比较分析,我们鉴定出共 1167 个基因在这两个阶段之间存在差异表达(其中孢子体上调 465 个,配子体上调 702 个)。

对这些世代偏向基因的功能进行了分析,我们运用了手动分配的功能类别体系,以及基于 GO(Gene Ontology)条目和 KEGG(Kyoto Encyclopedia of Genes and Genomes)通路的分析。
这些基因在两个手动分配的类别中表现出明显富集,分别为“细胞壁和细胞外”以及“细胞调节和信号传导”,此外还涵盖了一些未知功能的基因。

值得进一步注意的是,在这些世代偏向基因中,我们共鉴定出 23 个预测的转录因子以及 10 个 EsV-1-7 结构域家族成员。
后者在孢子体偏向基因组中表现出显著富集(χ² 检验 P = 0.001)。

有趣的是,孢子体偏向数据集和配子体偏向数据集均富含预测编码分泌蛋白的基因(Fisher's 精确测试 p = 2.02e-8 和 p = 4.14e-6)。
对与分泌蛋白相关的 GO 术语的分析表明,其富集模式与完整的世代偏向基因的情况相似,包括与信号传导、细胞壁和膜运输相关的术语,实验证明了人工指定功能类别的相对丰度,这些类别代表具有世代偏向基因的预测编码分泌蛋白。

根据前述分析,我们从差异表达基因列表中,筛选出了200个在孢子体和配子体世代间表达显著差异的基因,随后,对这200个基因的表达模式进行了进一步分析。
同时选择了奥罗和山姆突变体,以及第三个突变体,即立即直立(伊姆河)作为对照,这些突变体不会导致不同生存周期世代之间的转换。

从中可以观察到,奥罗或山姆的突变导致了配子体中某些基因的上调,同时孢子体中其他基因的下调,这与孢子体到配子体表型功能转变的趋势一致。
而且奥罗和山姆突变体呈现出相似的表达模式,并且在伊姆河变种人中,这种模式却显著不同,结合形态和生殖表型的信息,奥罗和山姆突变体的表现支持了ORO和SAM作为配子体向孢子体转化的主要调控因子。

这些结果为我们深入了解基因调控下的生命周期转换提供了关键线索,同时也强调了奥罗和山姆在该转换过程中的重要性。

 ORO和SAM蛋白在体外相互作用
ORO和SAM蛋白在体外相互作用作为生命周期调控因子或交配决定因子的HD TFs,通常会在异源二聚体复合物中进行构件的选择。
同时实验发现ORO和SAM蛋白也被证明能够在体外拉下法中形成稳定的异源二聚体。
缺失分析的结果表明,这两种蛋白之间的相互作用主要由它们的同源结构域介导。

 ORO 基因和 SAM 基因的进化起源和结构
ORO 基因和 SAM 基因的进化起源和结构经过序列数据库的详细分析表明,所有褐藻都拥有3个属于TALE类的HD转录因子,其中包括ORO和SAM的同源物,这些同源物在选择构建时具有共通性。
对褐藻中Oro和SAM同源物的比较揭示,Oro和SAM的HDs上游和下游区域表现出高度保守性,这些结构域不与已知的公共领域数据库中的任何结构域相对应,也不出现在公共序列数据库中的其他蛋白质中。
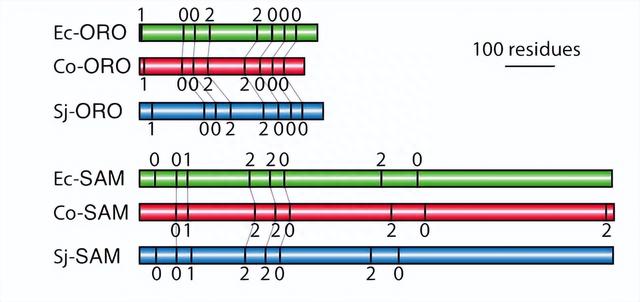
特别是,在与HD相关的区域中,我们未发现任何明显的相似性,尽管这些区域在真核生物中已被证实具有高度保守性。
但不能排除Oro和SAM蛋白可能具有这些结构域的不同变体的可能性,HD是Oro和SAM蛋白唯一共有的结构域。

为了识别Oro和SAM的远亲同源结构域,我们搜索了一系列具有特征性Oro和SAM蛋白结构域的滤过性HD-TFs,只有一种非褐藻蛋白,来自食草植物。
研究实验证明,eterosigma Akashiwo具有这些结构域的相似性,这使得暂时将其归类为ORO的同源基因,该菌株的转录组中还包括一个截断的TALE HD TF,其转录类似于SAM,但需要更完整的序列数据以确认与SAM的直系同源。

这一分析将ORO的起源追溯到与轮虫的共同祖先(约360 Mya),然而,ORO和SAM非HD区域的分歧排除了更远相关同源基因的检测。
通过进一步搜索Tale HD TF基因的内含子位置,发现这些位置与ORO和SAM没有检测到任何进一步的同源物

实际上,有学者认为,通过修改基于同源结构域的调控网络,在绿色谱系中可能在孢子体的复杂性产生中扮演了关键作用。
关键的事件可能包括替代真正的BEL类蛋白,以及Gsp 1类的BELL相关1基因,与真正的BEL类和KNOX类TALE HD TF之间的多样化替代。

特别值得注意的是,在链霉菌早期的进化过程中,两个KNOX亚家族的出现和亚功能化被认为促进了更为复杂的孢子体转录网络的进化。
在褐藻中,ORO和SAM也扮演了主要的发育调节器角色,然而,在这个谱系中,尽管多细胞孢子体出现,但并没有伴随着显著的TALE HD TF的扩大。

实验观察结果表明,新的机制已导致生存周期调节器的TALE HD TF的激活群体在复杂多细胞体中演化出,在褐藻(Ectocarpus siliculosus)中,谷氨酸受体GLR2被认为是这一机制的一部分。
这种现象并不意外,因为作为主要调控机构的招募TALE HD转录因子,应该与修改这些系统本身的调控机制相关。

对这些TALE HD TF的调控进行修饰可能会在生命周期中产生灵活性,例如,在褐藻中,如果要求严格的配子融合才能形成Oro-SAM异二聚体,那么在孤雌配子体中就不可能实施孢子体发育程序。


本研究的分析结果揭示了两个TALE HD TF蛋白质能够形成异源二聚体,同时对褐藻的生存周期中孢子体程序的部署具有关键作用。
在绿色植物谱系中,同样观察到TALE HD TF对孢子体程序的部署产生调节作用,这表明它们在生存周期调控中具有类似的功能。

尤其值得注意的是,KNOX类TALE HD TF基因的敲除,如MKN1和MKN6在Physcomitrella patens中导致孢子体世代转变为具有功能的配子体。
与金黄藻(Chlamydomonas reinhardtii)的胞子体到配子体的转变以及Ectocarpus中的Oro或Sam突变体观察到的表型相近,尽管这两个谱系已经在超过十亿年的演化中分离,它们仍然独立地演化出复杂的多细胞性。

这两个真核超群中生存周期调控因子的相似性表明,它们系源自一个共同的祖先系统,可以追溯到早期的真核生物进化。
这一生存周期调控系统的古老起源得到了其他事实的支持,比如在真菌和社会性阿米巴中,远亲同源结构域或类似结构域的蛋白质扮演交配类型因子的角色。

现在有越来越多的证据证明(HD TFs)在真核细胞生命树的bikont和unikont分支的生存周期调控中扮演着古老的角色,这些分支包括广泛的生物种类。
通过对这些独立、复杂、多细胞的谱系进行比较分析,我们能够识别出这些调控因子的约束情况,从而为深入理解复杂多细胞性产生的进化过程提供了重要的洞察。

一个特别引人关注的问题是,这些HD TFs是否也在协调动物生命周期的进展和发育中发挥作用?
本实验对于近亲TFs在动物单细胞中的功能的详细分析有助于更深入地探究这个问题。
引用[1]阿伦A,彼得斯,马克·考克,SM ( 2013 ) 模型中生命周期转变的非细胞自主调节褐藻 胞外 新植物学家 197 :503-510。
[2]布坎· ,明内西F,纽金特,布莱森·K,琼斯DT ( 2013 ) PSIPRED 蛋白质分析工作台可扩展的 Web 服务 核酸研究报告 41 :W349–W357.
[3]SM,戈德弗罗伊,阿伦A,勒考吉勒G,,旋塞JM (2011) OUROBOROS 褐藻配子体向孢子体生存周期转变的主要调节因子 Ectocarpus PNAS 108 :11518×11523。
