文:回溯档案
编辑:回溯档案
由于生物圈中固氮生物的长期进化,生物固氮(BNF)被认为是最环保、可持续的获取氮的方法之一,大豆作为人类植物蛋白的主要来源,在人类食物供应中发挥着不可或缺的作用。
促进大豆结瘤、增强全固氮被认为是最有效、最便捷的增产方法之一,近年来,纳米技术在农业中的应用逐渐引起人们的关注,纳米技术可能是农业可持续发展的潜在突破口。
通过功能性纳米产品增加豆类结瘤是一项突破性的尝试。

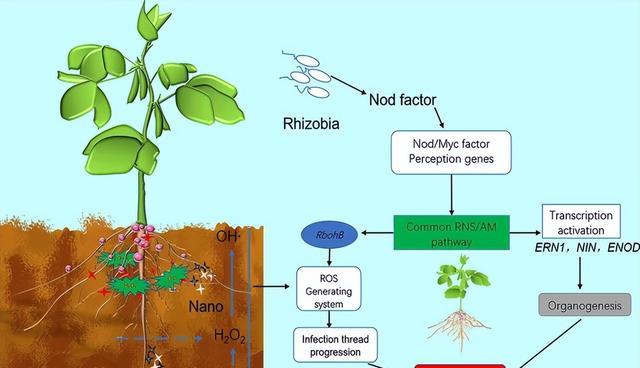
活性氧作为高活性氧衍生物,包括超氧阴离子、羟基自由基、氢过氧自由基和过氧化氢,这些自由基是在生物和非生物胁迫下的有氧代谢过程中产生的。

尽管具有毒性,ROS 仍可以作为信号来触发代谢调节以应对环境压力,ROS在豆科植物结瘤过程中在结瘤因子(NF)的诱导下产生,并参与感染形成,有助于细胞壁重建、基质糖蛋白(MGP)交联以及肌动蛋白微丝成核和分支。

ROS 通过空间和时间的改变与 Ca 2+一起调节结瘤。ROS 的共生生产主要通过细胞质膜 NADPH 氧化酶呼吸爆发氧化酶同源物 ( Rbohs ) 和Rbohs来完成过度表达刺激根瘤菌感染和根瘤形成,导致共生微粒体数量和固氮效率增加。

相反,抑制Rbohs会减少 ROS 的产生并下调根瘤菌诱导的过氧化物酶 ( RIP )、NIN和ENOD2的表达,总的来说,豆类中的 ROS 是一组调节结瘤的信号分子。

在结瘤过程中,为了控制过多结瘤对植物造成的胁迫,微小RNA作为反馈调节开关,可以通过剪切减少GmNNC1蛋白的量GmNNC1 mRNA,并减轻转录抑制子GmNNC1对GmENOD40的抑制作用,从而使GmEOND40进一步激活信号转导通路,最终启动结节的发育。

当根瘤过多时,根源性结瘤特异性CLAVATA/ESR相关(CLE)肽GmRIC1和GmRIC2通过芽源性抑制剂(SDI)如细胞分裂素诱导结瘤自动调节(AON)以抑制根瘤形成,miR172c的表达,从而避免大豆过度结瘤。

进一步的研究表明,一些材料,如锰基、钴基和铜基材料,在类似 FTR 方面表现出很高的效率,纳米材料介导的FTR也广泛应用于生物医学和环境保护领域。

Mn 2+和 H 2 O 2之间的类似 FTR 反应中生成 ?OH 的能力低于 H 2 O 2和 Fe 2+之间的 Fenton 反应,但前者(Mn 2+参与)可以延长ROS的诱导时间,在FTR体系中添加一定量的Mn 2+可以显着提高FTR效率。

据报道,Mn 2+与H 2 O 2反应可生成O 2 ?? 。FTR中产生的Fe 3+在O 2 ??的介导下可以被还原为Fe 2+,形成可持续的FTR 。
种植条件
将大豆种子在次氯酸钠中搅拌表面灭菌4分钟,用无菌水漂洗6次,转移至空白琼脂培养皿中,在黑暗培养箱中于25℃下培育发芽,将发芽的大豆种植在装有基质的无菌穴盘中,种植深度为3厘米。

定植后,每32株穴盘每三天浇灌2L无氮营养液,接下来,在温室中培养幼苗(14小时光照/10小时黑暗循环,光强度为150μmol m -2 s -2)。
根瘤菌接种
根瘤菌活化后,收集单菌落,接种到YMA液体培养基中,28℃,摇床(150rpm)培养72小时,备用。

然后用无菌水清洗根瘤菌,将菌液的吸光度值(OD 600 )调整为0.5。幼苗定植1周后,在每个植物根部周围的基质中接种1mL菌液。
采样时间
接种后第4天,采集根部样品,观察根毛的卷曲情况以及H 2 O 2和O 2 ??的染色情况,在根瘤菌接种后第 4、8 和 12 天对根进行取样,用于转录组测序分析。

第12、16、20天采集样本,观察结节发育状况,第28天采集样品评价根瘤重量、根瘤数、株高、根长等生长指标,根瘤完全成熟后(第28天),采集有根瘤的根系测定固氮酶活性,观察成熟根瘤切片,评价共生区的发育形态和根瘤菌的生物活性。

羟基自由基的产生
将纳米材料(浓度为1.25 mg mL -1)和离子材料(离子材料中的离子浓度与MF-NPs一致)分别制备成母液,配制成1 M亚甲蓝(MB)溶液。
随后,将100μL材料母液(纳米材料或离子材料)和200μL MB溶液与2mL 0.3%H 2 O 2溶液在振荡下混合,反应0、6、12、18、24、30、36、42、48小时后进行光谱扫描,测定OD 660进行定量分析。
H 2 O 2和 O 2的检测
如前所述,通过 DAB 和 NBT 染色对植物根部中的H 2 O 2和 O 2 ??进行可视化,通过用50mM Tris缓冲液(pH 5.0)中的0.1mg mL -1 DAB对根进行染色来分析根H 2 O 2含量。

O 2 ??含量是在含有 0.1 M NaCl (pH 6.1) 的 20 mM K-磷酸盐中与 2 mM NBT 一起孵育后测量的,根图像是在明场光下拍摄的。
大豆的转录组分析
在将根瘤菌接种到含有或不含10 mg L -1 MF-NP(作为CK)的蛭石培养基后第4、8和12天,从植物中收集整个根组织,立即放入Eppendorf管中,并储存在液氮。

使用植物RNA提取试剂盒提取总RNA。将获得的总RNA在Illumina NovaSeq测序平台上进行测序,构建mRNA测序文库,随后使用 hisat2 软件将测序读数映射到大豆基因组。

使用 DESeq 软件与P来鉴定差异表达基因(DEG)值 < 0.05,错误发现率 (FDR) < 0.05,并且 |log2 折叠变化| ≥ 1 作为阈值。
细菌活污渍和死污渍
采用冷冻切片机(Leica CM1950)进行显微切片,观察结节内的活菌和死菌,每个切片用SYTO 9和碘化丙啶(PI)染色剂的混合物染色,并在黑暗中保持30分钟,然后在荧光显微镜下观察。

细胞膜完整的细菌显示绿色荧光,而细胞膜受损的细菌则显示红色荧光。
统计分析
所有数据均以平均值±标准差表示,实验一式三份进行,通过独立样本t 检验确定组间的显着差异,并通过应用单向方差分析分析统计差异,显着性水平确定为 *P < 0.05、**P < 0.01 和 ***P < 0.001。
结瘤和固氮效率评价
结瘤始于类黄酮诱导的根瘤菌感染,导致结瘤因子(Nod-F)的表达,在结瘤过程中,ROS首先诱导根毛卷曲,然后诱导感染线和根瘤原基的发育,在这项研究中,我们计算了早期卷曲的根毛。

MF-NPs暴露增加了成熟区根毛的卷曲程度,是对照组的3.38倍,从而为成熟根瘤的发育和生长提供了先决条件,随着结节的进一步生长,形成肉眼可见的早期未成熟结节,根据其不同阶段的数量来定位结节的发育时期。

在20天的生长期内,观察到对照组和MF-NP治疗组之间未成熟结节分布的差异,20 d生长期未成熟结节数为对照组的7.43倍,表明暴露于10 mg·L -1MF-NPs可以延长结节的早期发育,导致结节数量持续增加。

通过根表型和根瘤发育来评估MF-NP处理对地下部分生长发育的影响,根部形态分析显示MF-NPs对根部没有明显的抑制作用,根瘤切片进一步分析显示感染区与非感染区的细胞形态和大小以及运输的发展没有明显差异。

组织正常SYTO 9和PI荧光染色显示,根瘤细胞中感染的根瘤菌表现出活力,MF-NPs处理组感染根瘤菌总数(绿色)和死亡根瘤菌(红色)数量与对照组相当。
对照组. 这些观察结果表明,MF-NPs暴露对大豆根系和根瘤的生长发育没有明显影响,甚至可以增加根长。
基因表达分析
在植物生长过程中,锰离子参与光合作用、呼吸作用、蛋白质合成和激素反应等过程),先前的研究表明,玉米(Zea mays)的生长仅在200 mg kg -1 Mn暴露下受到抑制。

而向日葵的生长直到Mn浓度达到5300 mg kg -1才受到抑制,理论上暴露0~100 mg L -1 MF-NPs不会对植物生长造成严重胁迫,因此大豆植株矮化并非由锰离子胁迫引起。
ROS 胁迫会影响植物根系和高度 ,因此本研究中大豆株高的变化可能是ROS调控所致,我们的数据表明,尽管株高显着降低,但大豆植株的总生物量却显着增加,这可归因于单株根瘤总固氮效率的提高。

然而,这种优势在暴露于高浓度的 MF-NPs 后消失,表明过高的 ROS 应激对总固氮有抑制作用,我们的元素组成分析表明,含有铁锰双离子的体系中生物量明显高于CK组,这与我们对铁锰离子之间FTR的假设一致。

尽管M-NPs+F-NPs的组合也含有铁离子和锰离子,但这种组合未能表现出促进作用,这可能是由于两种离子之间的物理距离所致,MF-NPs暴露组的根瘤数量和重量显着积累,这可能是由于ROS对大豆早期结瘤系统的正向调节作用所致。我们的结果与之前的一份报告一致。
结论
我们的研究结果表明,通过延长结瘤期可以有效增加根瘤数量,而不威胁植物的营养生长或触发结瘤自动调节(AON)途径。这项研究为诱导超常规结瘤提供了一种有效的策略。
本研究通过转录组学分析从基因水平和代谢途径证实了早期实验现象,以往的研究报道,碳纳米材料的诱导可以增加莲子早期结瘤基因的表达,但相关的作用机制尚不清楚。
我们的研究为探索这一机制提供了证据,工程纳米材料 (ENM)(Ag、ZnO 和 TiO 2)对豆科植物苜蓿A17的毒性作用研究,发现早期结瘤基因显着下调,ENM处理下结瘤的抑制可能主要是由于锌离子诱导的非生物胁迫。
在非豆科水稻中,荧光二氧化硅 (F-SiO 2 ) ENM 处理已被证明可以增加生物量积累,转录分析表明 F-SiO 2 ENM 处理可以上调与葡萄糖代谢和碳固定相关的基因的表达。
参考文献
【1】罗杰斯 C,奥德罗伊德 GE。工程谷物中氮共生的合成生物学方法。J 实验机器人。2014;65:1939–46。
【2】萨阿加、莫雷诺 YMF、卡西奥菲 BAM。植物蛋白作为人类饮食的优质营养源。食品科学技术趋势。2020;97:170–84。
【3】洛瑞 GV、阿维兰 A、吉尔伯特森 LM。农业技术革命中纳米技术的机遇和挑战。国家纳米技术。2019;14:517–22。
【4】Pulizzi F. 纳米农用化学品的兴起。国家纳米技术。2021;16:1056–1056。
【5】Kah M、Kookana RS、Gogos A、Bucheli TD。对纳米农药和纳米肥料与其传统类似物的严格评估。国家纳米技术。2018;13:677–84。
