
文:回溯档案
编辑:回溯档案
磷作为植物发育和生长所需的许多重要化合物的重要组成部分,积极参与光合作用、呼吸作用、碳水化合物代谢、能量转导等过程,然而,在许多农业系统中,土壤中可吸收磷的浓度不足以确保植物生产力。

作为最重要的粮食和经济作物之一,大豆是人类和动物食品的重要蛋白质来源,然而大豆的生长和产量受到多种土壤因素的限制,特别是土壤中磷的有效性较低。

低磷胁迫下,大豆植株矮化,叶面积减少,下部叶片可能出现坏死斑。此外,开花时间会增加,但在结果期间,豆荚数量可能会减少,由于这些磷介导的对大豆的表型影响,磷肥被用来克服不良的潜在产量结果。
大量磷肥容易固定在土壤中形成螯合物或沉淀物,可能造成环境污染,为了缓解缺磷对大豆生长的影响,寻找磷高效大豆材料、研究高磷高效基因显得尤为重要。
低磷胁迫诱导的四组主要基因,一组参与感应低磷信号,包括拟南芥初生根中发生的局部磷酸盐感应PHR1活性的长距离磷酸盐信号传导。

另一组成员通过涉及磷稳态的内部调节来介导磷酸盐分布,包括拟南芥中的SPX家族和包含SPX结构域的PHO1,其余两组基因主要集中于磷酸盐的获取和运输。

例如,GmACP1编码一种酸性磷酸酶,其在根中的过度表达可以增加营养液中磷酸盐的吸收,PHT1家族由大豆编码的磷酸盐转运蛋白中的15个PHT1基因组成,已被证明不仅参与指导根部磷酸盐的吸收。

而且还参与以磷酸盐形式运输磷,一般来说磷酸盐的获取和运输很大程度上取决于根系,因为在缺磷条件下根系形态会发生变化以维持植物生长。

这包括玉米侧根数量和长度的增加,耐低磷的大麦和拟南芥品种的根毛更长和数量更多,并且当低磷相关转录因子OsMYB2P-1过度表达时,水稻的初生根相对较长。

同时,与大豆磷效率相关的QTL定位也已取得进展,与种子磷含量、磷效率相关的根性状以及基于花和豆荚脱落率的大豆对低磷胁迫的耐受性相关的QTL已被确定此外。
通过对大豆高效磷材料的筛选,鉴定出高效磷基因型,如HP119、HP134、华夏1号、华夏2号等。

作为一种非生物胁迫,低磷条件可能引发一系列复杂的反应,研究表明植物激素信号通路对磷酸盐缺乏有强烈反应,当拟南芥和玉米遭受低磷条件时。

与植物激素信号传导相关的大量基因的表达会诱导或抑制根部生长,此外长期缺磷条件下,次生代谢产物对低磷胁迫的变化表现为花青素的积累。
在玉米中观察到类似的结果与苯丙素途径相关的基因被确定为在磷饥饿下上调或下调。

利用基因表达谱和转录组分析技术,在拟南芥、水稻和玉米中发现了大量与低磷胁迫相关的基因和信号通路,最近,曾通过高通量测序鉴定了大豆根中的磷酸盐缺乏反应基因。

选择代表性材料并考虑不同组织将进一步揭示与低磷胁迫相关的基因和途径,本研究通过水培实验,从219份大豆材料中筛选出耐低磷的春豆和低磷敏感的云和丰窝豆。

然后对这两个品种的根和叶进行微阵列芯片分析,通过分析基因表达数据,我们确定了42个候选基因和3个低磷胁迫诱导的常见途径。
本研究为高磷高效基因的功能鉴定提供了候选位点,对培育高效磷大豆种质可能具有重要意义。

低磷耐受种质CD和低磷敏感种质YH的鉴定
通过对219个大豆种质在不同磷处理条件下持续10天的水培实验,我们发现大豆种质CD和YH在+P处理条件下表现出相似的根系形态,同时表现出明显的根系形态。

-P处理条件下的形态差异,然后我们将种质CD鉴定为低磷耐受种质,将种质YH鉴定为低磷敏感种质。
因此,0.5mmol/LP和0.005mmol/LP的半Hoagland分别被认为是正常和低磷处理条件,CD中的初生根长度比YH中的初生根长度长,CD的根毛和侧根数量远多于YH,对CD和YH的根和叶进行了微阵列实验。

不同磷处理条件下表达差异显着的基因
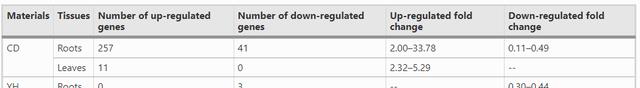
低磷条件下,耐低磷材料CD的根和叶中分别发现257个和11个上调的差异表达基因;与低磷处理相比,低磷敏感材料YH的根和叶中没有发现DEGs上调。
对于下调的DEG,在CD的根中仅发现41个DEG,在根中发现3个DEG,在YH的叶中发现7个DEG。

根中的DEG数量多于叶片中的数量,有趣的是在CD的根和叶中都发现了一种常见的DEG,它编码一种基质金属蛋白酶,倍数变化分别为4.10和3.59。先前的研究表明,它与大豆的病原体感染有关。

为了评估两个大豆品种中低磷胁迫和正常条件之间DEG的潜在功能,将基因本体论类别应用于低磷诱导的基因,这三个GO术语被分类为生物过程、分子功能和细胞成分。

我们发现DEGs与创伤反应、茉莉酸刺激反应、氧化还原酶活性和细胞壁显着相关,无论它们是上调基因还是下调基因,此外我们利用KEGG确定了与低磷诱导基因相关的途径。

总体而言,P值小于0.05的45条路径是在低P压力下激发的,大多数途径与代谢过程有关,值得注意的是,最重要的3条富集途径包括苯丙素生物合成、甲烷代谢和苯丙氨酸代谢,均与次级代谢产物的代谢相关。

不同大豆种质间表达差异显着的基因
种质CD和YH在承受低P胁迫的能力方面表现出巨大差异,CD(309)中的DEG比YH(10)中的DEG多得多,为了进一步揭示CD和YH之间与低磷条件相关的DEG,我们比较了大豆材料CD和YH之间的DEG。

我们对这195个标记的DEG进行了GO富集和KEGG分析。GO分析表明这些基因主要参与次生代谢过程,例如类黄酮生物合成。它们还参与对激素刺激以及防御和非生物刺激的反应。

此外它们还参与电子载体活性和过氧化物酶,KEGG分析显示富集的通路大多与次生代谢产物的代谢相关,包括苯丙素生物合成、甲烷代谢、苯丙氨酸代谢、黄酮类生物合成等。

不同磷处理条件下的317个DEG被认为是响应低磷胁迫的基因,不同大豆品种中的195个标记DEG不仅是由低磷胁迫诱导的,而且还由取决于对低磷条件的响应的材料变化诱导。

因此,我们比较了317个DEG和195个标记基因,发现85个重叠基因被认为是低磷胁迫诱导的活性基因,进一步的研究表明,大多数常见基因集中在根部,这与不同磷条件下根部的DEG数量多于叶子的情况一致。

根和叶之间显着差异表达的基因
在我们的研究中,在低磷处理下,根中检测到的DEGs数量多于叶片,表明根部在幼苗期存在更多响应低磷胁迫的基因,发现根和叶之间的差异表达基因。

低磷条件下诱导的DEGs根据其在正常条件下的表达模式分为三类,对于低磷胁迫下CD根和叶之间的6370个上调基因,三类DEG为5137个上调基因、1个下调基因和1232个在正常条件下根/叶中没有表现出差异表达的基因。

在1232个基因中,有97个基因在低磷条件下根和叶之间的表达差异大于4倍。此外,我们发现63、19和73个基因在低磷条件下分别与1207、893和1075个基因表现出大于4倍的表达差异,通过1个相反表达的基因,根和叶之间有253个标记的DEG。

我们对这253个标记的DEG进行了GO富集和KEGG分析,GO分析显示这些基因富含一组生物过程,例如氧化还原、氧化应激反应和非生物刺激反应,KEGG分析显示,大多数这些基因富含甲烷代谢、苯丙素生物合成和苯丙氨酸代谢。

低磷引起大豆的系统性变化
磷对大豆生长发育很重要,为了检测低磷胁迫相关基因,进行了多项比较分析,首先我们比较了给定材料和组织中不同磷处理之间的表达水平变化,这代表了应对低磷条件的材料内和组织内DEG。

同时,我们分析了不同磷浓度下材料间和组织间的基因表达模式,最后,我们获得了材料间和材料内、组织间和组织内之间的重叠DEG,以及不同P处理、材料和组织之间的共同基因。
通过这些比较分析,我们获得了响应低磷胁迫的稳定基因,并获得了大豆低磷胁迫相关的共同基因和通路。

在大豆根部磷缺乏响应基因的GO富集分析中揭示了17个生物过程,在这些过程中,我们的研究还检测到了光合作用、铁离子转运、脂肪酸代谢过程和应激反应。

通过对GO富集结果的分析,我们发现氧化反应以及过氧化物酶被反复检测到,早期研究表明,过氧化物酶的表达水平与对生物和非生物胁迫的抵抗力密切相关,如盐胁迫、创伤、疾病、过敏反应等。

最近的研究证实,在限磷条件下,在玉米、水稻和拟南芥中观察到包括过氧化物酶基因在内的氧化应激相关基因的改变。

结论
磷饥饿导致大豆基因表达的系统性变化,在这里通过微阵列芯片在低磷耐受性和低磷敏感性种质的根和叶上调查了低磷胁迫下大豆的基因表达模式。
通过对差异表达基因的比较分析,我们确定了42个候选基因和3个与低磷胁迫高度相关的常见途径,包括甲烷代谢、苯丙氨酸代谢和苯丙素生物合成。
这些结果不仅促进了我们对磷缺乏反应的分子基础的理解,而且促进了改善大豆磷利用率和设计高磷效率大豆的研究,这一过程可以优化肥料的使用并促进可持续农业实践的发展。
参开文献
【1】拉戈萨马两合公司。磷酸盐的获取。年鉴植物生理学植物分子生物学。1999;50:665–93。
【2】沙赫特曼DP、里德RJ、艾林SM。植物对磷的吸收:从土壤到细胞。植物生理学。1998;116(2):447-53。
【3】BureauM,MederskiH,EvansC。磷肥原料和土壤磷水平对大豆产量和磷吸收的影响。阿格伦J.1953;45:150–4。
【4】豪厄尔RW. 大豆的磷营养. 植物生理学。1954;29(5):477–83。
【5】曹敏,佟志,韩敏,程浩。磷高效利用大豆菌株的选育与评价。农作物。2001;4:22-4。
