
文字/编辑:一口半夏
«——【 ·前言· 】——»
丛枝菌根真菌(AMF)在耕种土壤中普遍存在,与主要农作物的根部形成共生关系,研究表明在受控条件下的AMF 具有促进寄主植物生长的潜力。
然而该领域的实际效益却很难估计,尤其是因为缺乏合适的AMF控制。

在这里我们实施了一种新策略,将 AMF 抗性选择性纳入遗传图谱群体中,以评估玉米对 AMF 的反应。
本研究的目的是为了推测出AMF约占谷物产量多大的比例范围,才会影响到不同植物基因型的相对表现,从而推导出对应的实际收益。

根据宿主反应的遗传结构表征,我们确信菌根依赖性和效益之间存在权衡,为玉米种植效益提供更好的方案。
 共生基因CASTOR和POLLUX
共生基因CASTOR和POLLUX为了产生用于田间实验的抗 AMF 玉米品种,我们鉴定了共同共生基因CASTOR和POLLUX ( DMI1 ) 的玉米(变种 B73)直向同源物。

与水稻一样,CASTOR和POLLUX在玉米中被发现是单拷贝基因,并命名为ZmCastor (GRMZM2G099160; Zm00001d012863; 染色体 5, 1 Mb) 和ZmPollux (GRMZM2G110897; Zm00001d042694; 染色体 3, 177 Mb)。
通过搜索公开的遗传资源,确定了两个突变子(Mu) Castor基因中的转座子插入:castor-1(翻译起始位点上游 44 bp)和castor-2(翻译起始位点下游 39 bp)。

在用 AMF 进行控制接种后,我们看到了Castor-1植物的真菌定殖,但发现Castor-2植物没有根内部真菌结构,反映了两个等位基因中转座子插入的不同位点。
我们改进了castor-2(以下简称castor)突变体进行更详细的定量表征,发现突变体中被破坏的转录本,并确认在接种粗制(沙罐生产)或平板培养(体外生产)接种物后不存在真菌定植。
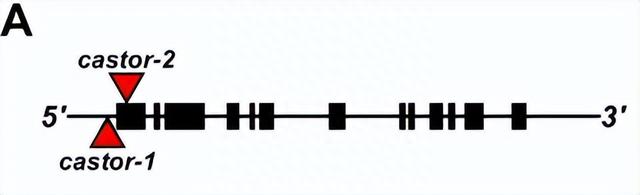
选定 AMF 的抗性来源后,我们开始生成群体,用于绘制现场宿主反应图,为此将原始温带W22 背景的蓖麻突变体与亚热带自交系 CML312 杂交,并进行两轮自花授粉以产生 F 2:3家族。
使用 PCR 基因分型来选择Castor基因座上野生型或突变等位基因纯合的F 2个体,这样所得的 F 3家族就突变而言是“固定的”。

通过这种方式,我们生成了 73 个纯合野生型(易感; AMF-S)和 64 个纯合蓖麻(抗性;AMF-R)的作图群体。
在Castor基因座之外,不同的家族根据 W22 和 CML312 基因含量进行了分离,使用全基因组标记对F 2亲本进行进一步基因分型,以构建用于 QTL 作图的遗传图谱。

数据证实了对AMF的抗性在田间是稳定的,并且在高营养田间条件下,蓖麻突变不会导致植物发育出现任何严重缺陷,也不会在菌根中表现出明显的多效性效应。
为了表征现场宿主对 AMF 反应的遗传结构,实验在雨养和中等投入条件下的重复试验中评估了 73 个AMF-S和 64 个AMF-R家族,选择了代表中等规模、亚热带生产的农艺情景,以及我们认为 AMF 可能发挥重要作用的条件。

总体而言AMF-R家族倾向于表现出轻度褪绿,并且与AMF-S家族相比,其耳朵明显较差。
同时随机抽样植物的根部,发现AMF-S家族中存在明显的 AMF 定植,而AMF-R家族中不存在 AM 真菌结构。

实验中为了估计总体宿主反应,还比较了所有耐药家族的易感者,通过收集了十五个物候和形态特征以及产量组成部分,发现AMF-S和AMF-R家族之间的 15 个性状中有 10 个存在显着差异,对应于 3% 到 51% 的阳性宿主反应(表 1); 此处的正定义为生长性状和产量成分的更大值或加速/更同步的开花)。

AMF-R家族身材矮小,吐丝延迟(雌性开花),导致开花期(雄性开花)-吐丝间隔延长——非生物胁迫的典型症状玉米。
AMF-R家族的穗大小以及穗和总粒重均减小,AMF-S和AMF-R系列之间的单个颗粒的重量或大小没有显着差异。
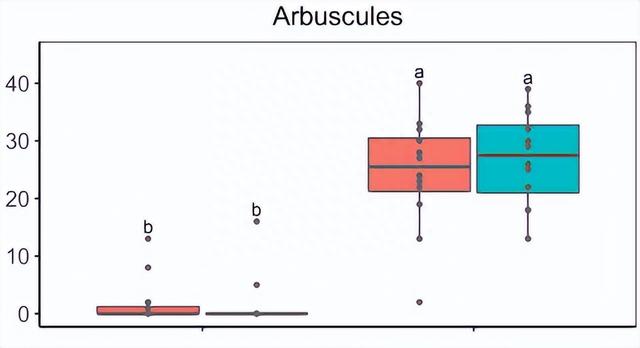
然而,AMF-R家族中每穗的籽粒总数减少,表明结实率较差,这可能是 ASI 增加的结果。
 菌根反应变化的QTL x AMF 效应
菌根反应变化的QTL x AMF 效应使用 F 2:3家族获得的性状估计值及其各自 F 2亲本的基因型进行了数量性状位点 (QTL) 分析,在一次分析中结合了AMF-S和AMF-R家族,包括Castor基因型作为交互协变量。
在此模型下,Castor加性效应(以下简称 AMF 效应)估计了所有家族的边际宿主反应,QTL 加性效应捕获了亲本之间的遗传差异,QTL × AMF 效应表明了 CML312 和 W22 亲本反应中潜在的遗传差异。

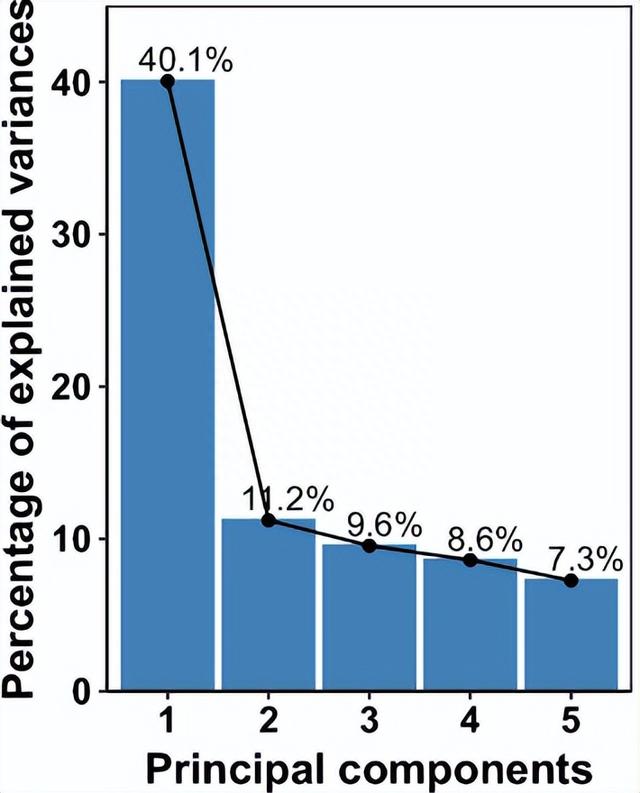
除了直接分析性状值外,我们还通过主成分(PC)分析组合了相关性状组,运行QTL作图中的前五个PC,并且鉴定了 28 个 QTL,其中 21 个显示 AMF × QTL 相互作用的证据。
这时将显着的 QTL、AMF 和 AMF x QTL 术语组合成每个性状的单个多 QTL 模型,并计算由 AMF、加性 QTL 和 AMF × QTL 术语解释的表型变异的百分比。
对于株高、穗重和总粒重,AMF 和 AMF x QTL 效应的组合解释了一半以上的总遗传变异(基于广义遗传力 H 2 的估计),以及超过四分之一的遗传变异。

显着 AMF × QTL 效应的鉴定揭示了 CML312 和 W22 亲本之间对 AMF 反应的遗传差异。
宿主反应的变化混淆了依赖性和效益的差异前者是给定品种在缺乏的情况下执行的能力AMF,后者表示工厂从该协会中获得利润的程度。
在确定了显着的 AMF × QTL 相互作用后,通过分别检查AMF-S和AMF-R家族中的 QTL 效应来区分依赖性和效益。

对于 17 个 QTL 中的 12 个显示出显着的相互作用,该效应是有条件的——主要表现在AMF-S或AMF-R家族,但不属于两者。
因此认为AMF-R家族中特异表达的条件 QTL代表依赖性变异,AMF-S家族中特异表达的条件 QTL是效益的变异。
基因组的两个区域与多个 QTL 相关,表明有一个共同的机制基础:染色体长臂 (chr) 上的一个区域与耳性状相关,而第四染色体上的一个区域与植物高度和开花时间相关。

使用 PC 组合性状进一步细化了这两个 QTL“热点”,证明了依赖性和效益之间的差异。
 拮抗QTL效应
拮抗QTL效应几个 QTL 显示出更极端的 QTL × AMF,其中该效应在AMF-S和AMF-R家族中都有表达,但符号发生了变化(即亲本等位基因的相对表现的“交换”)——描述的情况作为拮抗性多效性。
在这种情况下,菌根植物中的优势等位基因在没有菌根的情况下对性能有害,反之亦然,为单基因座水平的权衡提供了证据。
值得注意的是,几个与关键产量成分相关的 QTL 显示出拮抗性多效性,我们在整个基因型水平上观察到类似的“等级改变”效应。
尽管作图设计不允许直接评估与AMF-S和AMF-R相同的家族,但可以将 QTL 模型拟合到 AMF 两个级别的所有基因型,由此产生的估计显示了有或没有 AMF 的情况下性能排名变化的证据。
为了进一步探索基因型水平的反应,我们使用全基因组模型来预测 137 个科的菌根和非菌根性状值。
具体来说是使用了AMF-S家庭训练一个菌根性能模型,并将其应用于整个人群,同样使用AMF-R家族来训练非菌根性能模型。
全基因组预测与训练集的观察值非常一致,尽管它们没有捕获极端观察值,这两个模型的比较表明,在一种条件下与主要产量成分最高值相关的基因型,在另一种条件下则无异常或较差,这表明依赖性和效益之间存在权衡。

 结论
结论数据表明AMF共生对中等投入、雨养亚热带田地的玉米性能做出重大贡献,将我们的观察结果推断为每公顷种植 80,000 株植物的耕地(试验地点当地的典型情况),估计 AMF 的定植将贡献约2吨/公顷,总产量约为 5.5 吨/公顷。
虽然粗略的数字需要谨慎对待,但这一估计可以表明 AMF 的重要性。

我们在此描述的方法还可用于评估旨在促进或丰富土壤健康的做法的功能影响,包括整体“有机方案”或具体干预措施,例如减少耕作或应用外源 AMF 或其他微生物。
当在低至中等养分利用率下生长时,自交玉米品种始终表现出对 AMF 的积极反应,在其他作物物种中AMF 的影响更加多变,从互利共生到寄生都是连续的,在高粱(玉米的近亲)中,宿主反应从积极到消极变化,具体取决于植物品种和 AMF 物种。

然而宿主反应通常是在受控条件下生长的幼苗中测量的,获得的值可能低估了AMF共生在田间的真正重要性。
就玉米的具体情况而言,重要的是要将大多数已发表的报告中所描述的自交系与栽培中使用的杂交品种和开放授粉的地方品种区分开来,杂种优势往往会减少依赖性,从而减少宿主对 AMF 的反应。
这并不意味着丛枝菌根在杂交玉米栽培中不重要,只是比例反应可能随近交程度的不同而不同。

实验中F 2:3家族分离了约 50% 的基因组,任何给定个体都保留约 25% 的杂合性,因此该材料介于 F 1杂种和近交系之间。
将蓖麻突变基因渗入不同的遗传背景将能够产生AMF-R杂种,并随后评估其在田间试验中的表现,如果有合适的突变体,我们的方法可能很容易应用于其他作物。
分析表明宿主依赖性和获益之间存在功能性权衡:在 AM 易感家族中预测为最佳的基因型在 AM 抗性背景下被预测为不显着或较差,反之亦然。

在个体基因座水平上,与吐丝天数、穗重、总粒重、每行粒数和五十粒重相关的 QTL 出现了拮抗多效性的例子。。
例如增加根皮层通气组织(根空气空间)的比例可以减少根系的碳需求,提高觅食效率,但代价是限制 AMF 的定殖以及潜在的宿主反应。

令人有些惊讶的是,形态性状(例如雄穗分支数量)的 QTL 取决于对 AMF 的敏感性,表明多效性系统信号也在发挥作用,可事实上在植物发育(包括雄穗结构模式)中,发挥关键作用的相同植物激素也参与AMF共生的建立和调节。
例如乙烯信号传导的破坏已被证明可以调节烟草中 AMF 的(负)宿主反应水平;赤霉酸信号通路中的 DELLA 阻遏蛋白对于 AMF 在水稻中的定植至关重要;涉及独脚金内酯的反馈回路可调节 AMF 在蒺藜苜蓿中的定植程度。

重要的是许多相同的信号已成为驯化和植物改良的目标,在许多情况下,QTL × AMF 效应是由与温带 W22 等位基因相关的可塑性驱动的,
除了影响产量之外,我们观察到的遗传结构还表明 AMF 在影响产量稳定性方面发挥着作用。

但鉴于 AMF群落在不同地点或年份的不可预测性,植物宿主的优化可能需要更多的时间。
引用[1] 奥热·RM、托勒 (2015) 丛枝菌根共生在干旱条件下比在充足浇水条件下更能改变寄主植物的气孔导度:一项荟萃分析
[2]本德SF、范德海登 MGA (2016) 一场地下革命:生物多样性和土壤生态工程促进农业可持续发展
[3] 邦方特·P (2015) 丛枝菌根对话:你说的是“植物”还是“真菌”?
