
文字/编辑:一口半夏
«——【 ·前言· 】——»
黄二铁蛋白(FDPs)是一类存在于细菌、古细菌以及真核生物中的普遍存在的复合酶,值得注意的是,藻类Anabaena sp. PCC 6803生物中涵盖有四种FDP蛋白(Flv1-4),这种构成在光合作用过程中的光保护机制中扮演着不可或缺的角色。
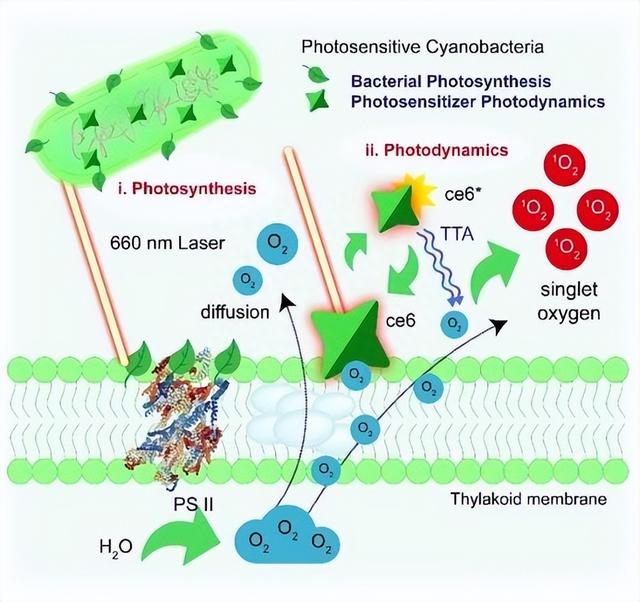
本研究中,我们直接比较2高浓度(3% CO)和低浓度(空气中的一氧化碳)下光致氧的还原情况,即类梅勒反应(2,HC)和无机碳条件(LC),主要为了鉴定出,Flv1/Flv3杂化二聚体是维持稳态氧(O)还原的唯一有效机制。

实验最终目的是为了在低浓度环境的情况下,通过分析强烈诱导的NDH-1配合物与电子之间的相互竞争,从而进一步确定Flv1/Flv3在瞬态电子汇方面所发挥的功能。

 LC和HC条件下类梅勒反应
LC和HC条件下类梅勒反应本研究我们采用了膜进样质谱(MIMS)结合18富氧技术,在光照条件下区分光合总氧和O2的产生,同时对18O2和16O2的吸收进行了深入探究。

在实验条件下通过光合氧化体系,第二至第四的氧化还原体系细胞,在低碳浓度(LC)和高碳浓度(HC)环境中,均积累了大量的Flv2、Sll0218和Flv4,并在空气中(CO2浓度大于1%)以及高碳浓度环境(HC)条件下,证明细胞中更高的O2光还原速率。

在低碳浓度(LC)和高碳浓度(HC)条件下,flv4-2/ OE(过表达)细胞与野生型(WT)细胞的Flv 3蛋白水平相似,这强有力地支持在光合过程中,flv4-2操纵子蛋白对O2光还原在细胞内的贡献。
值得注意的是,在LC条件下生长的flv4-2/ OE和WT细胞的O2进化率之间没有显著差异,然而,在flv4-2/ OE细胞中,总O2进化率显著增加。

如前所述,二氧化碳浓度(Ci)水平对于FDP在转录本和蛋白质水平上的表达产生显著影响:Flv2、Flv4和Flv3的表达已在低碳浓度(LC)条件下得到证实,并在转向高碳浓度(HC)条件时表达。
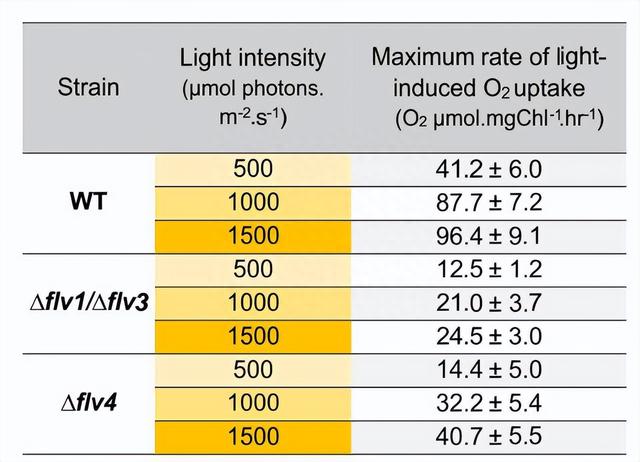
从黑暗环境中的转变后,野生型(WT)细胞在低碳浓度(LC)和高碳浓度(HC)条件下,呈现出显著快速的光诱导O2摄取,具体地说,其摄取速率分别为每小时59±6.4和56±6.4微摩尔氧化2分子/毫克叶绿素a。

这个迅速的响应阶段在本实验中得到了表现,然而,在低碳浓度(LC)条件下培养的细胞与在高碳浓度(HC)条件下培养的细胞相比,其O2光还原动力学表现出差异。
当野生型(wt)细胞的培养经过快速诱导期{i}后,明显地观察到了O2相的猝灭现象。

在O2还原过程中,首先经历了一个强烈的衰减相,其持续时间约为1分钟,随后,进入了一个相对稳定的状态,在这个状态下,O2还原速率约为每小时33 ± 5.9微摩尔氧化2分子/毫克叶绿素a,这一过程发生在照明期间。
与此相反,在 HC 生长的 WT 细胞中,光诱导的氧还原速率在阶段{ I }中仅在最初的2-3分钟内略有下降(从约56 ± 2微摩尔氧化2分子/毫克叶绿素a)。

在实验过程中我们发现,O2光还原动力学在flv4-2细胞中与其相应的野生型(WT)细胞类似,在HC条件下,光还原动力学趋于稳定,而在LC条件下,flv4-2细胞的O2光还原反应则呈现出强烈的双相猝灭现象。

为了研究在LC条件下的吸收和呼吸与O2光还原之间的关系,我们进行了针对M55突变体(Δ ndhB)的实验,M55突变体由于其疏水性的NdhB亚基缺失,因而缺乏整个NDH-1复合物。
在LC条件(pH 8.2)下生长的M55突变细胞表现出与野生型类似的特性:在光诱导条件下迅速产生O2光还原(第一阶段),随后维持稳态状态,但缺乏O2的第二阶段现象。

综上所述,在光合暗适应阶段转向光照环境后的光还原过程中,M55突变体的行为与野生型(WT)细胞存在差异,同时与野生型细胞相比,M55突变体表现出较为缓慢的诱导过程,并且总氧进化速率明显降低。
这提示M55突变体的稳态O2光还原状态并非源于从PSII增加的电子流。

 生长介质pH值和碳酸盐浓度对梅勒样反应的影响
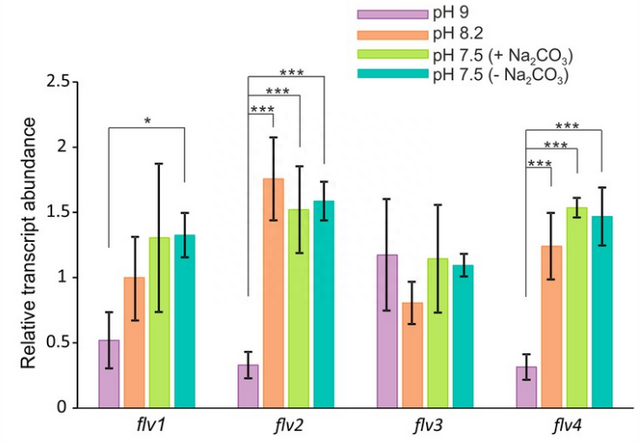
生长介质pH值和碳酸盐浓度对梅勒样反应的影响我们对生长培养基的pH值以及其中碳酸盐的存在进行了评估,以探究它们可能作为调控剂影响梅勒样反应的程度和动力学,以及在(LC)条件下FDPs的积累情况。
在pH值为9的环境中生长的WT细胞表现出明显而短暂的Mehler样反应特征:O2光还原率在最初的30秒光照后达到最大值,然后在接下来的1分钟内迅速降至初始暗态的氧水平。

与WT相似,flv4突变体细胞在光照条件下也只表现出短暂的O2光还原特征,然而,在pH9的环境中生长的flv1/∆flv3和∆flv3/∆flv4突变体细胞中,并没有明显观察到O2光还原的迹象。
特异性抗体免疫印迹的结果表明,在高碳浓度条件下生长的WT细胞,在低碳浓度且pH值为9的条件下,几乎无法检测到Flv2和Flv4蛋白的存在。

与蛋白质数据的一致性相符,对于pH为9的细胞环境,我们观察到Flv2和Flv4的转录水平均显著下调,这强烈暗示了pH对于四联体Flv2和Flv4的转录调控存在重要的依赖性。

这些结果极具说服力地表明,在pH为9的条件下,明显且短暂的O2光还原在野生型(WT)和flv4突变细胞中,主要归因于Flv1/Flv3异二聚体的活性。
通过上述数据,我们能够对O2的不同动力学阶段的起源作出初步结论
 在PSII和PQ池水平上,FDP所诱导的氧离子还原过程
在PSII和PQ池水平上,FDP所诱导的氧离子还原过程为了进一步确定Flv2/Flv4异二聚体相关的O2光还原过程,我们将重心放在flv4-2/OE突变体身上,其在LC、pH值为7.5以及不含碳酸盐的条件下进行生长。

在这个突变体中,我们观察到Flv2和Flv4蛋白的积累显著增加,同时其O2光还原速率较WT更为显著,当线性电子传递受到细胞色素b6f(Cyt b6f)水平的限制,尤其在使用DBMIB作为抑制剂的情况下,这种现象更为显著。

在使用DBMIB作为抑制剂的情况下,flv4-2/OE突变细胞表现出强烈的光诱导O2吸收,与之预期一致,在存在DBMIB的情况下,Δcyd突变细胞的光诱导O2吸收未被检测到。
 光强度变动对FDP缺失突变体生长表型的影响
光强度变动对FDP缺失突变体生长表型的影响通过实验证明,Flv 1/flv 3杂二聚体通过在Mehler类反应中作为一种有效的电子汇发挥作用,使细胞在波动光下生长。
然而,目前的研究结果清楚地表明,在Mehler样反应中的Flv2/Flv4异二聚体的额外参与,特别是在LC的条件下和在pH值为8.2或更低中选择所需的构件。

这些发现使我们更准确地研究了生长培养基的pH值,和波动生长光条件(FL)对各种FDP突变体生长性能的综合影响,为达此目的,我们施加了严格的光照条件,FL20/500)和温和的光照条件(FL50/500)来模拟光强度的波动。

在严格的条件下,每隔5分钟会暂停20 µ 摩尔光子m −2 s −1 背景光,借着30秒光脉冲强度为500摩尔光子m ~ 2 s ~ 1 与 温和( FL50 / 500 ,当 50 个 { mol 光子 m - 2 秒 - 1 个背景光,每 5 分钟被 30s 光脉冲强度为 500 的脉冲干扰时) μ mol 光子 为 2 s 为 1 波动的光施加在不同的pH值水平。

从实验结果来看Flv1/∆flv3突变体(∆flv3/∆flv4)在严峻的光波动条件(FL20/500)下无法维持生长,而这一现象与生长介质的pH值无关。
然而,在较为温和的光波动条件(FL50/500)下,这一突变体在碱性pH值(例如pH 9)环境下的生长速度较野生型相比较慢,这一现象同样在pH 8.24的条件下观察到。

同时实验研究发现Flv4突变体在所有不同的pH水平下,无论是在轻度或重度光波动条件下,都表现出与野生型细胞相似的生长特性。
更值得注意的是,在pH值为7.5和8.2的条件下,第二阶段∆sll0218和第四至第二层OE突变体的生长情况也得到了研究。

 Flv 2/Flv 4杂二聚体在LC条件下Mehler反应
Flv 2/Flv 4杂二聚体在LC条件下Mehler反应通过对不同科囊藻突变体中各种FDP积累的特异性影响进行表征,我们得以阐明Flv2、Flv4以及Flv1、Flv3蛋白在体内O2光还原过程中的参与。
人们普遍认为Flv1/Flv3蛋白在HC和LC条件下保护PSI,编码的蛋白质,而由 flv4-2 在LC下高度表达的操纵子在PSII的光保护中起着重要作用。

通过系统的实验和分析发现, 科囊肿 表达的Flv4 大肠杆菌 能够产生依赖于NADH的O 2 -体外还原作用,然而与来自厌氧原生动物的FDP的活性相比,报告的反应速率极低(几乎是残留的)该酶对NADPH无亲和力
同时科囊肿 蛋白质导致一种说法,即Flv3在依赖NADH的O2光还原过程中,与二聚体可能发挥相似的作用。
与Flv1-OE和Flv∆3-OE(或Flv1∆3-OE)突变体相比,Flv2蛋白的亲和力降低了(对NADPH的亲和力非常低),这些结果清楚地表明,在体内的光还原过程中,Flv3(或Flv1)的同聚物并不在O2中发挥作用。
尽管O2的光还原能力通常与科囊肿的FDP丰度相关,然而蛋白质丰度并不是决定O2光还原能力的唯一因素。

实际上,尽管高效且稳定的O2光还原在高浓度(HC)条件下的细胞中得以观察,但却几乎无法检测到Flv 2和Flv 4的水平,这与低浓度(LC)条件下的水平相比,Flv 3的含量较低。
此外,通过在pH 7.5条件下省略从BG-11生长培养基中获取的碳酸钠,我们观察到O2光还原率的增加,并且这并没有引起FDPs的显著转录和蛋白质水平的变化,这提示了可能存在氧化还原调节酶的活性。


在本研究中,我们提供了关于Flv2/Flv4对体内O2光还原的贡献的有力证据,通过应用光还原18O标记氧和实时气体交换测量不同FDP缺失突变体。
同时也深入分析了其中的细节:灭活Flv2或Flv4被证明导致O2光还原的幅度显著减少,相较于野生型细胞,而过表达Flv4-2操纵子则将O2光还原速率提高了近两倍。

值得注意的是,小蛋白质 Sll0218 促进类似 Mehler 反应的可能性被排除在外,同时科囊藻的O2光还原能力通常与FDP的丰度相关,然而,蛋白质的丰度并不是决定O2光还原能力的唯一因素。
尽管如此,实验的数据并不排除Flv 2/Flv 2和/或Flv 4/Flv 4同齐聚物,也参与O以外的过程的可能性,这种情况发生在Flv 1和Flv 3蛋白中,它们作为同配体在波动的光照条件下对细胞进行光保护。

在pH 8.2的条件下,完全消除了光诱导的O2吸收现象,而在pH 7.5以及存在电子传输抑制剂DBMIB和HQNO的情况下,对于生长在此条件下的WT细胞而言,FDP驱动的O2光还原有所减少。
这显示FDP驱动的O2光还原并不在PSII或PQ-pool水平上发生(无论是通过Flv1/Flv3还是通过Flv2/Flv4)。

这个结论也得到以下事实的支持:与缺陷的FDPs的WT和突变体不同,cyd突变体在存在二溴甲基溴的条件下不表现出光诱导的O2吸收。
引用[1]阿拉维迪耶娃· ,大湖J ,张平 ,阿罗 ,( 2015 ) 黄二铁蛋白保护蓝藻的产氧光合作用
[2]巴特奇科瓦N ,失落JP ,飞行 N ,Keranen M ,卡梅尔D ,阿罗, (2010) Synechocystis 6803蛋白质组群的动态变化对定量蛋白质组学 揭示的CO(2)限制的响应
[3]卡西耶-肖瓦C ,肖瓦特F ,( 2014 ) 铁氧化物在蓝藻中的作用和调节, 集胞 藻PCC6803: 近期进展
