文:回溯档案
编辑:回溯档案
棉花是最重要的天然纤维作物,支持数十亿美元的生产和加工业,棉花属(棉属)由大约45个二倍体物种组成,属于八个基因组群(A-G和K)和5个异体四倍体(AD)物种。
在棉属中,五个四倍体物种中的两个(2n = 4x = 52;G. barbadense L. 和 G. hirsutum L.),以及两种二倍体物种(2n = 2x = 26;G. arboreum L.和G. herbaceum L.),在过去的几千年里,在新旧世界被独立驯化用于棉纤维生产。

在这四种栽培品种中,四倍体品种G. hirsutum,也被称为“陆地棉花”,约占全球棉花产量的95%。因此,世界上绝大多数棉花育种计划都集中在改良陆地棉花上。
随着全球对纺织产品的需求不断增加,来自合成纤维的激烈竞争以及纺织工业通过转向高速纺纱技术的现代化,对具有更高产量和改善纤维质量的陆地棉花品种的需求从未如此重要。

然而产量增加通常与陆地棉的纤维质量呈负相关,产量与纤维质量之间的这种负相关阻碍了棉花育种工作,以改善多种性状。
棉花育种者一直非常希望有效地打破这种负面联系。随机交配程序提供了一种重要的方法来打破不良关联并在几种作物中形成新的组合,包括烟草,高粱,大豆和燕麦。

以前,随机交配也被证明可以降低棉花性状之间的相关性,随机交配需要花费大量的时间和精力,如果从一大群不同的亲本系开始,它提供了一个打破不利连锁块并形成新的重组的机会,其中一些应该是优越的。
传统的棉花育种计划,主要依赖于杂交适应基因型和基于表型选择新的等位基因组合,在过去的一个世纪中为棉花工业的成功做出了巨大贡献。

然而,如上所述,产量和纤维品质性状的非常复杂的定量遗传以及它们之间的负关联要求棉花育种者制定更有效的策略来开发优良品种。

标记辅助选择(MAS)就是这样一种策略,在过去三十年中受到植物和动物育种者的高度关注,在棉花中已经发表了许多关于绘制定性和定量性状的报告。
至于绘制与纤维质量相关的数量性状位点(QTL),到目前为止已经报道了1000多个QTL,然而,这些QTL中的大多数是基于对种间(主要是在G. hirsutum和G. barbadense之间)种群的分析而获得的。

在这项研究中,我们首先开发了一个550个品系的重组近交种群,这些种群来自六个随机交配周期,从11个陆地棉花品种的半等位基因杂交开始,然后是六代自花授粉。
在两年内种植了550个重组自交系(RIL),以获得纤维质量测量,其次我们筛选了15538个简单序列重复(SSR)标记,以识别11个亲本中的多态性标记。

11位父母的遗传多样性
我们在15538个亲本中筛选了11个SSR标记的多态性,在这些标志物中,2132个(13.72%)是多态性的,当比较任何两个父母时,差异最大的一对是Acala Ultima和M240RNR(1231个标记,或7.92%),差异最小的一对是FM966和STV474(710个标记或4.56%)。

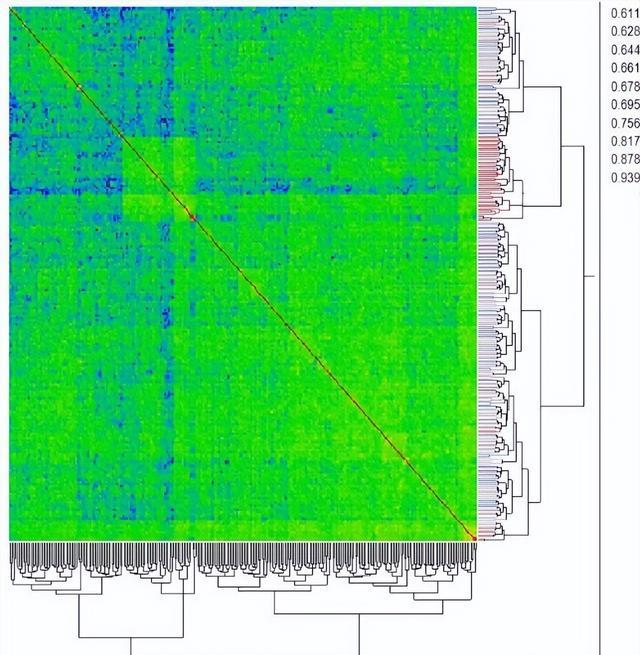
提供了使用42个基于 DICE 系数的 SSR 标记数据构建的 UPGMA 树状图树,尽管选择这11个亲本来代表美国棉花品种的广泛多样性,但整体遗传多样性非常低。
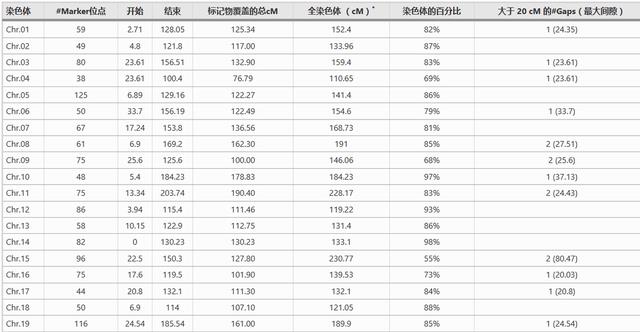
1582个多态性SSR标记的统计和基因组分布
根据高密度共识图谱,在覆盖了80%以上四倍体棉花基因组的43个多态性标记中,我们选择了1582个标记,这1582个标记显示,在1585个亲本中,共有100个显性位点、884个显性位点和11个单态性位点。
共鉴定出4483个等位基因,1582个选定标记的基因组分布,如果基于高密度共识图,这些标记覆盖了棉花基因组的约83%。

然而,如果基于G. raimondii基因组序列,则所选标记覆盖740 Mbp,占93.6%的覆盖率,由于A基因组参考序列不可用,本报告以四倍体棉花的遗传图谱为主要参考。
覆盖率不均匀,从55号染色体(Chr.)的15%到Chr.98的26%不等,间隙大于23 cM的间隙有20个,其中19个发生在17条染色体的端粒区域。
Chr.15的覆盖率最低(55%),在Chr.80的长臂上有一个47.15 cM的大间隙,没有发现单个多态标记,其可能原因将在后面讨论。

母体和 RIL 的纤维质量测量
11个亲本和RIL在美国密西西比州一式三份种植,RIL及其母体的纤维特性,在RIL中观察到所有性状的范围很广,并且存在违规隔离。

父母性状与RIL之间的皮尔逊相关系数比较显示在附加文件5中,RIL的相关系数通常低于亲本,这表明随机交配确实在一定程度上破坏了联系。
在RIL中,父母伸长率(ELO)和微米(MIC)之间的强正相关被打破,然而,在RIL中观察到束强度(STR)与上半部分平均纤维长度(UHM)之间存在显着的正相关。

QTL 集群
在与纤维品质性状相关的157个标记位点中,有37个位点与一个以上的性状相关。2、3、4和5分别与6、6、16和0066个性状相关。没有一个基因座与所有2个性状相关。

除了MIC是一个复杂的性状(纤维成熟度和细度的组合)外,每个QTL簇对性状的影响通常是相似的,换句话说,从育种的角度来看,通常能够增加 ELO、STR、UHM 或 UI 值但降低 SFC 值的 QTL 集群被认为是有利的 QTL。
不利的QTL聚类通常会增加SFC值,但会降低所有其他四个性状的值,值得一提的是,Chr.0114上的QTL簇(标记位点CMa)对这<>个性状(SFC、STR、UHM、UI)的有利育种效果最大。

相反,Chr.上的QTL簇(标记位点Ca)对SFC、STR和UI的不良繁殖影响最大。
种内遗传多样性和标记覆盖率
虽然本研究中使用的11个亲本品种在农艺性能和育种谱系方面具有相当多样的差异,但15538个SSR标记揭示的亲本品种之间的遗传多样性较低。

这些SSR标记是由世界各地的许多小组开发的,标记的来源要么来自基因组DNA要么来自许多基因型的EST序列,用于筛选的15538 SSR引物对未预先选择。
并且有理由相信这些标记涵盖了Wang报告的四倍体棉花的整个基因组,本研究进一步证实,栽培的旱地棉花具有非常狭窄的遗传多样性和密切的亲缘关系,这可能是因为在驯化过程中发生的一些瓶颈。

此外,在23个亲本中,有20个大于11 cM的基因组区域未发现多态性标记,这些区域可能表明现代陆地棉花中的固定(纯合子),并且可能包含通过育种和选择过程固定的关键适应性基因。
固定的纯合基因组块是构建中高密度种内多毛G.连锁图谱的巨大障碍,然而这些区域也可能不包含重要的农艺基因或QTL。

对这些区域的靶向测序可以更好地了解其生物学功能,值得注意的是,这23个基因组块中有17个发生在46条染色体的端粒区域。
这种现象引起的生物学影响尚不清楚,它们对实际育种的影响也不清楚,Gore在使用源自TM-24016和NM1之间杂交的RIL时观察到了相同的现象。

在我们之前对Ligon-lintless (Li 1)基因,我们揭示了类似的现象,蔡也是如此,Chr.15在其长臂末端有一个非常大的间隙(80.47 cM)。
在检查了共识图后,我们注意到150至230 cM之间的区域几乎完全用RFLP标记映射,同样,Chr.0的20和15 cM之间的区域也由共识图中的RFLP标记组成,Chr.15的总遗传距离很可能不像Blenda那样长到230 cM。

我们的标记覆盖率可能大于55%。事实上,我们的标记覆盖了G. raimondii Chr.43(对应于G. hirsutum的Chr.364或Chr.62)的598,467 bp和02,01,15 bp之间的范围,约占整个G. raimondii Chr.99的7.02%。Chr.06、23、24和25也存在类似的情况。

结论
棉花育种者在使用MAS改善纤维质量等数量性状时面临的主要挑战之一是,在不影响其他性状的情况下,很难将QTL从其他物种(如巴巴登斯)转移到陆地棉花中,虽然在种内种群中鉴定的QTL在育种中实施的障碍较少,但旱地棉花的低遗传多样性为研究人员提供了另一个挑战,即准确识别和绘制具有中度至高度影响的稳定QTL。
在这项研究中,我们使用涉及11个不同陆地棉花品种的随机交配重组自交系种群来鉴定纤维 QTL,这种方法确保了更高的多态性,使我们能够用1582个多态性SSR标记对RIL群体进行基因分型,这是迄今为止最多的。
再加上更大的种群(550 RIL,陆地棉花有史以来报告的最大单一种群),我们已经确定了131个纤维 QTL 和37个QTL 簇,p < 0.01。54个QTL被绘制在与文献中先前确定的QTL相同或相似的位置,而7个是新的。
使用这两个位点可以同时改善<>个纤维性状。我们将继续在一系列棉花品种中评估这些QTL,此外,我们正在多个地点测试这些RIL,以获得产量数据,并评估随机交配对打破产量和纤维质量性状之间的负相关关系的影响。
参考文献
【1】Wendel JF,Cronn RC:多倍体和棉花的进化史。阿格隆。2003, 78: 139-186.
【2】Fryxell PA:棉属L.(Malvaceae)的修订分类学解释。瑞德亚。1992, 2: 108-165.
【3】Meredith WR:MD 52ne高纤维优质棉种质和复发亲本MD 90ne的注册。 作物科学. 2005, 45: 806-807.10.2135/作物科学2005.0806.
【4】梅雷迪思·MR:定量遗传学。棉花。编辑:Koh JW,Lewis CF. 1984,美国威斯康星州麦迪逊:美国作物科学科学,132-147。
【5】Ulloa M,Meredith WR:种内人群中农艺和纤维质量性状的遗传连锁图谱和QTL分析。棉花学报 2000, 4: 161-170.
