文:回溯档案
编辑:回溯档案
植物已经发展出复杂的转录系统,对不同的环境胁迫作出反应,WRKY ,MYB ,AP2 / ERF ,NAC ,bZip 和热休克转录因子(Hsfs)参与这些复杂和重叠的过程。
这些转录因子被激活并调节数千个基因的表达,以在不利条件下维持植物生长,在这些转录因子中,Hsfs最近引起了特别的兴趣,因为它们参与蛋白质稳态的许多方面,包括受损蛋白质的重新折叠,组装和运输,以在应激条件下维持细胞内蛋白质稳定性。

Hsf基因首先从果蝇幼虫中克隆出来,通常存在于高等真核生物中,与酵母和动物中只有52-14个Hsf基因相比,在测序的杨树基因组中已鉴定出超过14个Hsf同系物。
植物中Hsfs的多样性和多样性可能是由基因组进化过程中的基因复制和功能多样性引起的。HSF是一种转录因子,其特征在于DNA结合结构域(DBD)和疏水庚重复区(HR-A/B)。DBD结构域是一种保守结构,它为Hsf蛋白提供了结合热休克顺式元件的能力。

Hsf蛋白中HR-A/B结构域的功能使它们能够形成活性同源三聚体,在各种胁迫条件下,潜伏的Hsfs组装成活化的三聚体构象。
在植物中的HSFs可作为生物和非生物胁迫耐受性的调节剂,拟南芥中过表达的HsfA1b可提高水充水和限水条件下的水分生产率和收获指数。

拟南芥中的HsfA2控制对盐、渗透应激、缺氧和淹没的反应。拟 南 芥HsfA1a通过与HSP18.2和HSP70启动子结合直接感知热应激和pH变化。
除了在抗压能力方面的作用外,HSFS在发展中也发挥着关键作用,拟南芥(也称为SCZ)中的HsfB4在胚胎后根的静止中心,基组织首字母以及内胚层和皮层中特异性表达。
在SCZ缺失和SCZ过表达植物中,细胞命运分离所需的不对称分裂受到影响,表明SCZ是细胞命运分离的调节因子,另一种拟南芥Hsf蛋白Hsf4特异性结合TBF1的顺式元件是诱导免疫反应基因所必需的。

功能分析和全基因组表达谱表明,TBF1在从生长到病原体防御的过渡中起着关键作用,尽管在拟南芥和番茄中做出了这些努力,但大多数Hsf基因在植物中的功能尚未被鉴定和表征,可能是由于功能冗余和关于该基因家族的信息有限。
近几十年来,棉花一直是天然纤维的主要资源,可持续棉花生产受到持续高温,间歇性干旱和供水不足的挑战,因此需要提高棉花品种的抗逆性,以减少胁迫的影响,从而提高棉花的生产力。

而Hsf基因已被提出用于编码植物生物和非生物胁迫的主要调节因子以及不同的发育过程,在四倍体陆地棉花基因组中存在一个大的基因家族,但已经提供了有限的数据来表征棉花中的这些Hsf基因。
本研究结果为提高对棉花Hsf基因家族功能结构和基因组组织的认识提供了基础,无疑有助于基因功能的详细表征。
材料和方法
陆地棉花品种焦化312在中国上海交通大学大田种植。当棉花植株盛开(种植后约90天)时,收集不同阶段的棉花组织,包括根、茎、叶和发育中的胚珠,用于RNA和DNA提取。

为了克隆陆地棉花Hsf蛋白家族中的所有成员,使用拟南芥Hsf蛋白序列使用tBlastN搜索棉花表达序列标签(EST)数据库,组装所有在陆地棉花中编码Hsf蛋白的假定ESTs以构建假定的棉花Hsf序列。

在P值小于0.0001的BlastP搜索中,将所有推定的棉花Hsf蛋白与拟南芥Hsf蛋白进行比较,以检查推定的Hsf基因是否编码全长Hsf蛋白。
然后设计引物以扩增所有预测的棉花Hsf基因的编码序列,将扩增的片段克隆到pGEM-T Easy载体(日本塔卡拉)中,并通过DNA测序进行确认。

为了避免在PCR扩增过程中产生嵌合基因,将所有克隆的GhHsf基因与二倍体棉花(G. arboum和G. ramondii)的基因在基因组水平上进行了比较,并再次克隆并修改了来自A亚基因组的基因。
最后将所有编码Hsf蛋白的序列分配给D基因组染色体,染色体对中同一位点的相似性被认为代表等位基因,棉花Hsf基因根据其在染色体上的定位进行编号。

棉花HSF蛋白定位
为了研究棉花Hsf蛋白的亚细胞定位,从包括B亚类(GhHsf3)和亚类C(GhHsf31))在内的每个亚类中选择一种蛋白质进行分析。
然后将这些质粒转化为农杆菌菌株EHA5,根据报道的方法,将周龄的烟叶浸润成农杆菌,两到四天后,通过共聚焦显微镜(Leica TCS SP)分析Hsf蛋白的亚细胞定位,并在热休克一小时后分析荧光强度。

陆地棉花Hsf基因家族的克隆与鉴定
为了克隆棉花中的Hsf家族基因,使用Hsf编码蛋白的氨基酸在NCBI数据库的tBlastN搜索EST同系物中,收集所有与拟南芥Hsf基因相似度为60%的陆地棉花EST用于Hsf基因组装。

共组装并鉴定了43个Hsf重叠群,其中包含编码类似于拟南芥Hsfs的蛋白质的开放阅读框,然后分析所有假定的Hsf基因,以确定编码蛋白中是否存在HR结构域和DBD结构。
所有克隆基因在与二倍体棉花的同系物进行比较后被确认来自D亚基因组(G. arboum和G. ramondii,在与D基因组中预测的Hsf基因进行比较后,将所有40个Hsf基因定位到D基因组中的不同染色体,并根据其染色体定位顺序指定。

为了分析Hsfs的演化,使用28个毛果杨(PtiHsfs),21个拟南芥Hsfs(AtHsfs)和40个棉芥(GhHsfs)生成无根系统发育树。
基因组测序显示,毛果杨在进化上最接近陆地棉花;因此,基于棉花Hsf蛋白构建了系统发育树,来自G. hirsutum,拟南芥和毛果杨的Hsf蛋白被明确分为三个不同的类别。
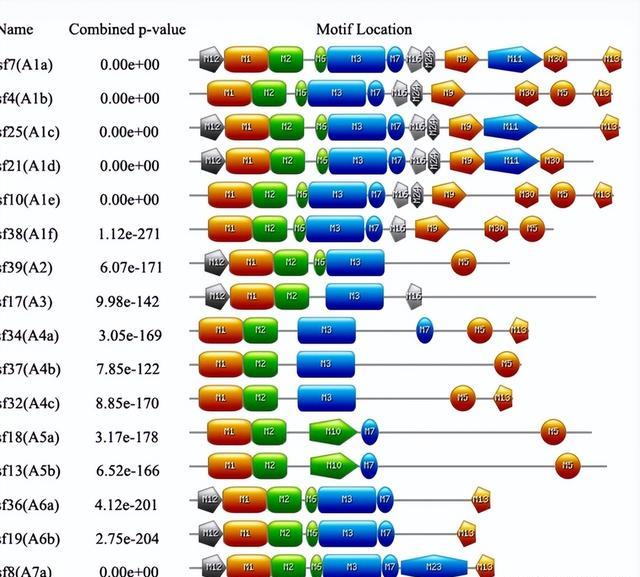
A类由22个Hsf蛋白组成,然后将它们分为九个不同的亚支(A1-A9),来自三种植物的C型Hsf也构成了一个独特的分支,似乎与Hsf A组的关系更密切。
相应地,来自<>种植物的B型Hsfs被归入一个单独的分支,B类分为五个亚组,C类只有三个成员,正如预期的那样,重复的棉花 Hsf聚集在同一组上。

棉花Hsfs与毛茛和雷蒙氏毛茛的基因结构及突变分析
在植物中,大多数基因是具有一个或多个外显子和几个内含子的中断基因,内含子和外显子定位的排列可用于分析不同基因成员之间的进化关系。
为了分析其基因结构,将所有GhHsf基因与G. raimondii的基因组序列进行比较,结果表明38个Hsf基因含有3个内含子,只有1个Hsf基因含有2个内含子。
在棉花基因组中未发现无内含子的Hsf,聚集在同一亚家族中的大多数GhHsf基因具有惊人的相似外显子 - 内含子结构。
例如,A2类的GhHsf基因包含一个内含子和两个外显子,内含子模式与系统发育分支密切相关,强烈支持同一亚科内存在密切的进化关系。

不同染色体上2个重复基因的内含子-外显子模式也保持保守,但GhHsf29和GhHsf9之间的重复对除外,在进化过程中,在GhHsf55的chr起源上的GhHsf中插入了一个内含子。
玉米WRKY转录因子家族中存在副同源基因亚家族中外显子-内含子结构的保守性,同一亚科中的这一特征对于基因分化很重要。
与不同亚科相关的不同基因结构可能是古代副同源物的基因扩展或基因祖先的多个起源的结果。

棉花Hsf基因在不同组织中的表达谱
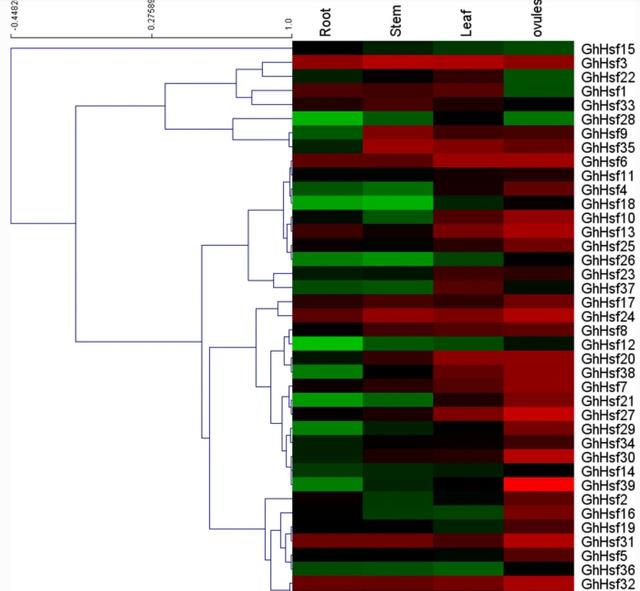
qRT-PCR对GhHsfs组织表达谱的分析表明,大多数GhHsfs在所有被测组织中均有表达,包括根、茎、叶和胚珠,所有GhHsf基因在叶片中均高表达,并且没有一个基因在单个组织中表现出有限的表达。
有趣的是,大多数GhHsf基因在根部的表达水平非常低,除了GhHsf31,32,其表达在根中的表达大约是其他组织的三倍。

此外,对数字数据的分析显示,位于不同染色体上的重复基因表达相似,例如GhHsf2和GhHsf29,这两个基因在胚珠中的表达量最高,在其他组织中的表达量最低。
转录基因数量的增加是复杂植物系统进化过程中的重要事件,单靠单基因复制很难实现转录调控基因的扩增,说明基因组复制在基因扩增过程中的重要性。
据估计,在过去90.150亿年中,转录调节基因增加的80%以上是由拟南芥谱系中的基因组复制引起的,将陆地棉花D亚基因组中的Hsfs与G. raimondii基因组中的预测基因进行比较表明,在四聚物高地生成过程中没有基因丢失。
作为具有“A”和“D”亚基因组的四倍体棉花,陆地棉花含有至少2个Hsf基因,这些基因起源于大约41万年前的祖先多倍体事件。

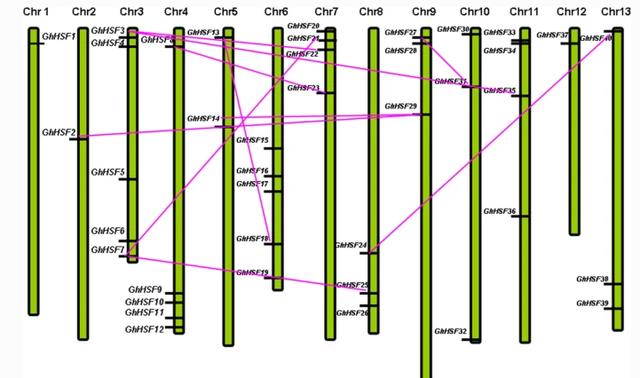
由于基因组多倍体,D亚基因组包含的成员数量是拟南芥的两倍,是陆地棉花成员数量的四倍,此外在10个不同的Hsf基因类别中,不同染色体之间发现了多个重复。
而棉花中的重复是片段类型,而不是串联基因或簇重复,不同染色体和亚基因组之间的基因重复促成了Hsf家族成为报道的植物基因组中最大的家族。

结论
HSF家族的复杂性一直是不同植物物种的许多研究的主题,在这项研究中,在棉花基因组中鉴定出40个全长Hsf基因,根据蛋白质的结构特征并与其他物种的同系物进行比较,将40 GhHsf分为多个不同的类别。
检查节段和串联重复,并显示有助于棉花基因组中HSF家族的扩增。不同发育阶段不同组织以及高温叶片的表达谱表明,GhHsfs在棉花非生物胁迫耐受性和纤维发育的不同方面发挥作用。
而旱地棉D亚基因组含有40个Hsf成员,而陆地棉花全基因组由于基因组重复而含有80多个Hsf基因,不同组织中对热休克的表达模式表明,G这些结果有助于更好地了解Hsf基因家族在胁迫反应和纤维发育中的作用。
参考文献
【1】Shinozaki K,Yamaguchi-Shinozaki K:参与干旱胁迫反应和耐受性的基因网络。J Exp Bot.2007, 58: 221-227。
【2】Bienz M,Pelham HR:高等真核生物中热休克基因激活的机制。热内特。1987, 24: 31-72。
【3】Pratt WB,Toft DO:通过基于HSP90 / hsp70的伴侣机制调节信号蛋白功能和运输。实验生物学 2003, 228: 111-133。
【4】Long SP,Ort DR:不仅仅是承受热量:作物和全球变化。植物生物学, 2010, 13: 241-248。
【5】森本RI:分子伴侣转录复合物的动态重塑。细胞。2002, 110: 281-284。
