导语
多倍体(基因组复制)是进化的关键力量。然而,多倍体细胞核中亲本基因组之间的相互作用,通常涉及亚基因组优势,知之甚少。竹子属于禾本科(Poaceae)中的单系群竹亚科(Bambusoideae),包含一个较小的草本以及一个二倍体的分支(126种)和三个主要的多倍体木本分支(1,576种)。先前的研究发现在木本竹子(WBs)中发生了两次独立的四倍体化事件,随后又发生了一次六倍体化事件,并涉及到三、四或五个灭绝的二倍体谱系的未解决假说。对于木本竹子,报告的染色体数目通常是恒定的(例如,四倍体的2n=(40)46-48,六倍体的2n=70-72)。这表明组成的亚基因组可能保持了未重组的状态。因此,竹子为研究古代多倍体起源植物的亚基因组优势进化提供了一个理想的系统。今天,小编就为大家解析这篇发表在《Nature Genetics》期刊上的有关竹亚科物种的高分多组学文章的研究思路~

文献标题:Genome assemblies of 11 bamboo species highlight diversification induced by dynamic subgenome dominance
发表期刊:Nature Genetics
影响因子:30.8(Q1)
发表时间:2024.03.15
研究思路

主要研究结果
1. 11个竹子的基因组序列分析
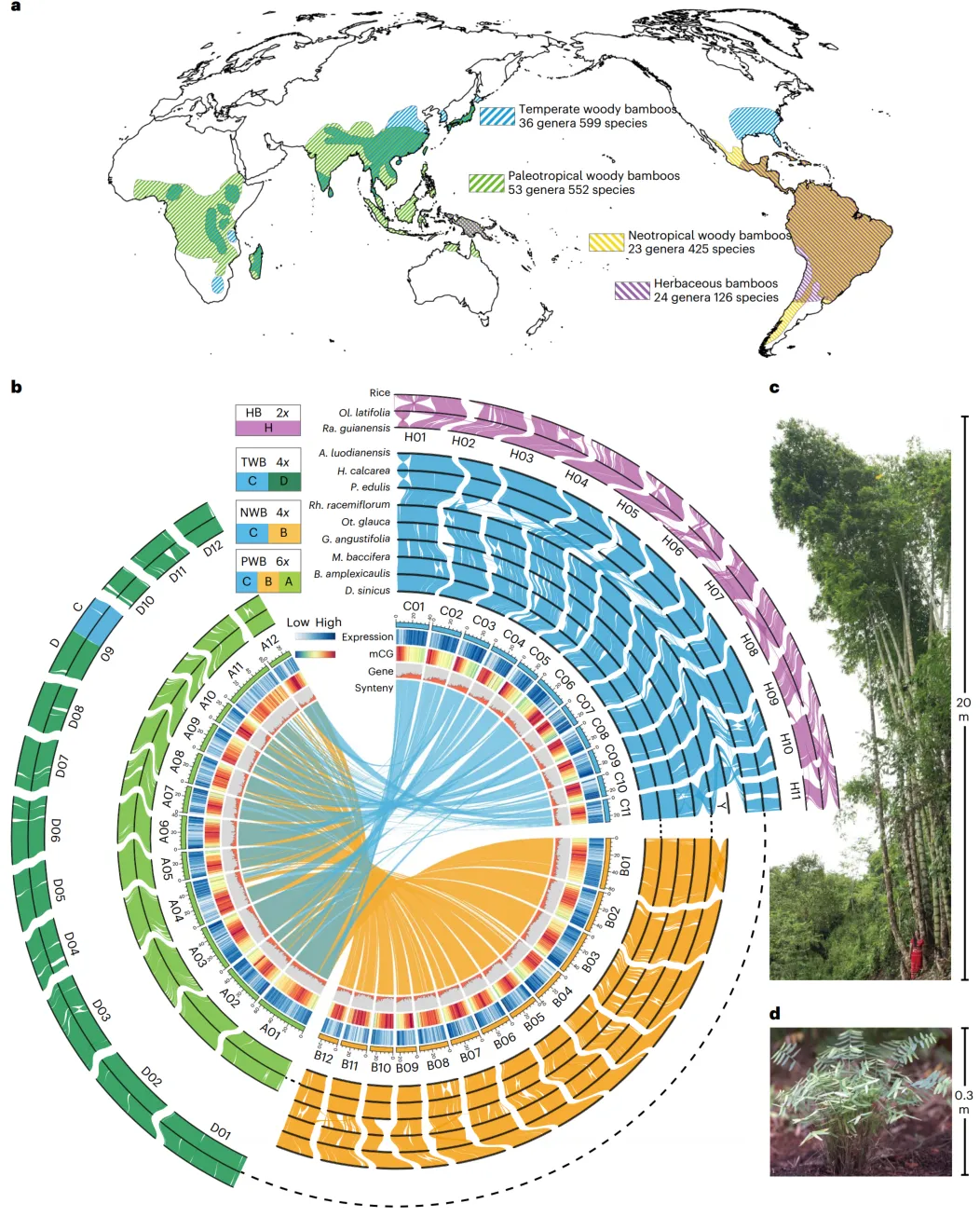
作为第三大的禾本科亚科,竹亚科在物种和形态上展现出极大的多样性。为了覆盖不同的倍性水平和系统发育多样性,选择了11种代表性物种进行基因组测序:两种草本竹(HBs,2x,Olyra latifolia和Raddia guianensis)和九种木本竹,分别属于三个类群:温带(TWBs,4x,Ampelocalamus luodianensis、Hsuehochloa calcarea和Phyllostachys edulis)、新热带(NWBs,4 x,Rhipidocladum racemiflorum、Otaea glauca和Guadua angustifolia)和古热带(PWBs,6 x,Melocanna baccifera、Bonia amplexicaulis和Dendrocalamus sinicus)。在这些物种中,巨龙竹D. sinicus是已知世界上最大的竹种,与草本竹Ra. guianensis形成鲜明对比。
结合平均为124.5x的Nanopore长读长和80.4x的短读长数据,这11个基因组被全新组装并校正为114至3,619个contig,平均N50分别为5.3 Mb和最大为17.5 Mb。利用染色质构象捕获(Hi-C)测序技术,来自这11个基因组的序列平均94.1%被锚定并一致地组装成二倍体、四倍体和六倍体物种的11、24和35个伪染色体。
2. WBs的亚基因组起源和多倍体化史
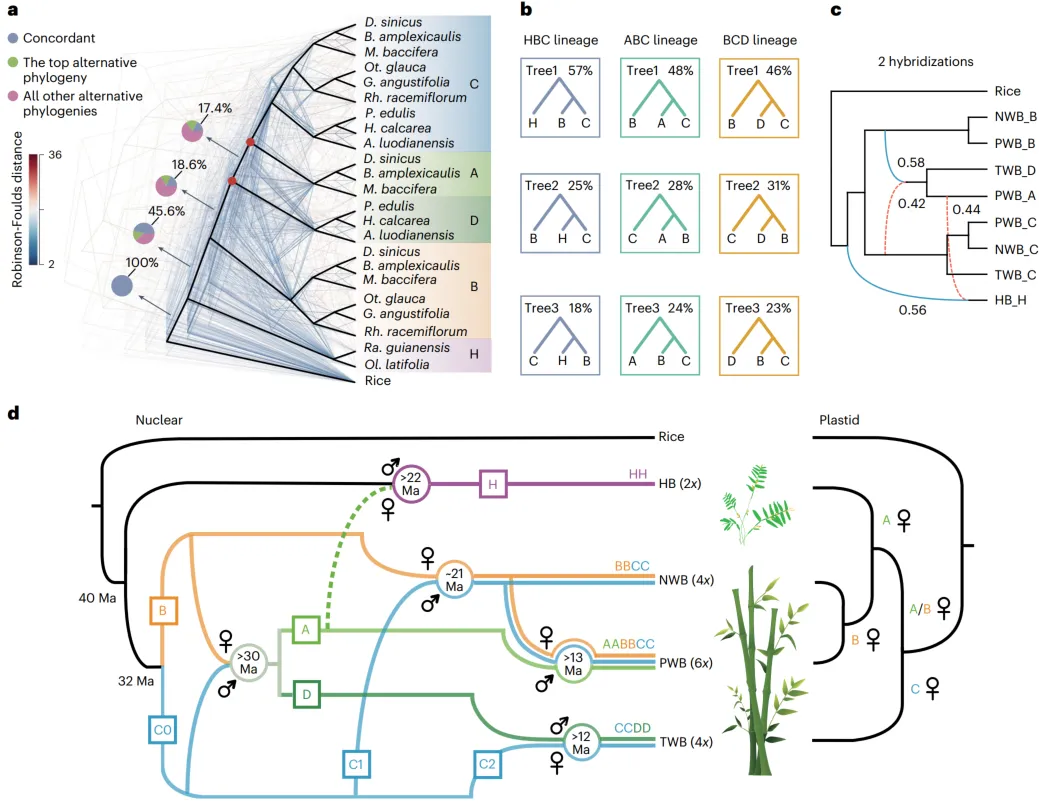
采用基于系统发育和序列相似性的策略对竹子的亚基因组进行了鉴定。收集了两个syntenic基因数据集,即456个“完美拷贝”syntenic基因(在二倍体、四倍体和六倍体竹子中分别具有1:2:3的预期拷贝)和13891个“低拷贝”syntenic基因(等于或小于1:2:3的拷贝),广泛分布在所有染色体上,用于系统发育分析。四个不同的WBs亚基因组,即A、B、C和D亚基因组,以及先前确定的HBs的H亚基因组,在两个数据集的分析中一致得到支持。序列相似性分析也支持亚基因组的识别,其中亚基因组A和D聚集在一起。
在456个同源基因中去除了26个异常值,并恢复了木质竹亚基因组谱系的单系性。然而,在基因树和共同演变的基于合并的树之间存在广泛的拓扑不一致性。着重研究涉及H、A和D亲本的主要冲突,最常见的拓扑结构占据了57%、48%和46%的基因树。基于此,推断了木本竹系统发育网络和推定的渐渗事件,并确定了B和C祖先谱系之间的杂交,导致一个杂交的二倍体祖先分化为A和D亲本,与上述基因树的不一致模式相符。渐渗分析还表明了第二个网状化事件,涉及H和A祖先,大约有16%的基因树支持这种情况。然而,来自其他木本竹二倍体祖先到H亚基因组祖先的渐渗可能也发生了,特别是如果这些序列信号在演化时间内被稀释,仅有弱证据留存。叶绿体系统发育和约7%的核基因树也表明了草本竹和木本竹的祖先之间的古老杂交。
综上所述,提出了竹类起源和多倍化的精细模型。草本和木本谱系的分化发生在竹类演化的早期,随后是木本祖先分化为两个(B和C)而不是四个或五个二倍体祖先。A和D的二倍体祖先很可能是通过B和C祖先之间同源杂交物种形成的,时间为3200万至3000万年前,前者作为母本。B和C1谱系之间的杂交后发生了多倍化,导致了新热带竹类(BBCC)的出现。随着四倍体作为母本,也观察到了小麦和燕麦中,第二次多倍化发生在1300万年前之前,导致了古热带竹类(AABBCC)的出现。第三个事件,还涉及C谱系(C2),1200万年前之前导致了温带竹类(CCDD)的起源。
3. WBs进化中的核型稳定性
除了新热带竹类(NWBs)和古热带竹类(PWBs)C亚基因组中的chr12的裂变和融合成chr3、chr6和chr11外,自它们大约在3000万至3200万年前分化以来,四个木本竹亚基因组一直保持着与12条染色体的全基因组同源性。高水平的同源性也在源自共同多倍化事件的多个物种之间得以保留,至少在最近的多倍化事件发生之后1200万年。然而,在 Rh. racemiflorum 中,最短的染色体(Y,38.9 Mb)没有同源染色体,以及基因密度和表达水平低于其他染色体。对竹类祖先核型(ABKs)的重建还表明,木本竹亚基因组,特别是A、B和D,类似于祖先草本科的核型(AGK),在漫长的演化过程中保持着令人惊叹的进化稳定性。亚基因组之间的大规模重排仅发现在由chr9D和chr2C大片段(38.9–54.8 Mb)融合而成的镶嵌染色体上,这被三个温带竹类(TWB)物种共享,表明这种情况发生在物种分化之前。在木本竹类中,亲源交换也以0.43%至1.27%的低水平发生。相比之下,草本竹类(HBs)中发现了许多重排,包括chr10和chr12的融合和伴随的染色体数目减少。

4. WBs的亚基因组优势和转变
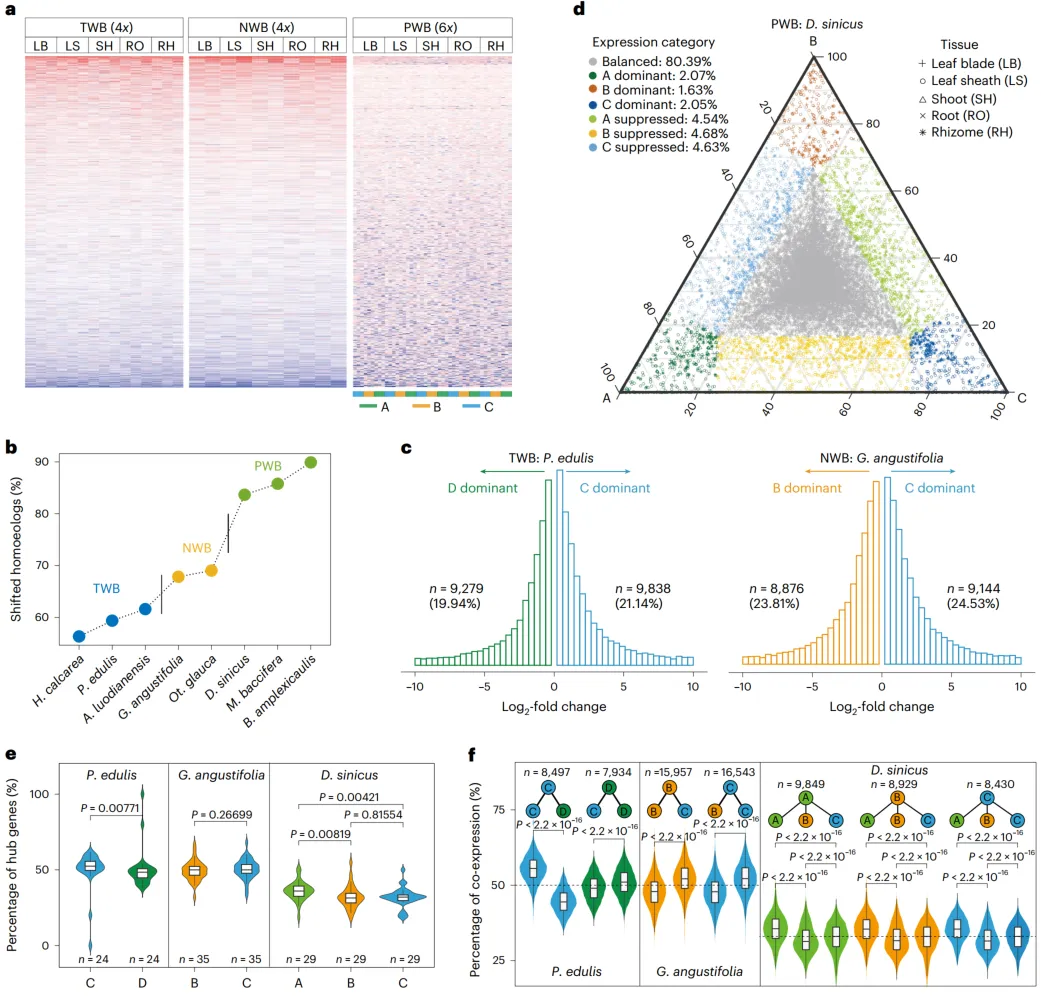
为了捕获多倍化后转录组景观的变化,对11个测序的竹类进行了476个转录组样本的测序和分析,代表不同组织在各种发育阶段,每个物种每个组织有三个生物学重复。在木本竹类中,与草本竹类相比,基因在组织间的表达广度较低,指向亚基因组表达差异。与木本竹类中的其他三个亚基因组相比,C亚基因组始终具有更高比例的表达基因,以及最高的平均表达水平。
为了确定每个类群中亚基因组的表达模式,鉴定了所有三个温带竹类(TWB)和新热带竹类(NWB)物种共享的4,123个和3,839个1:1同源基因对,以及古热带竹类(PWBs)的1,157个三联基因(1:1:1)。主成分分析(PCA)显示了表达在组织间的清晰分离(PC1和PC2),随后在所有三个类群中,亚基因组之间的清晰分离(PC2和PC3)。这种分离也观察到在具有更多同源基因的个体物种的分析中。在两个四倍体类群中,同源基因在组织和物种间的上调或下调表达模式保持一致,而在PWBs中变化很大,呈现出镶嵌状。根据它们的表达模式,同源基因进一步被聚类为10个组。超过一半的基因对(在TWBs中为58.5%–63.5%,在NWBs中为66.9%–68.1%)和大多数的三联基因(82.7%–88.9%)分化成不同的组。
研究总结
该研究展示了对竹类系统(禾本科:竹亚科)的分析,该系统包括从二倍体(草本)到四倍体和六倍体(木本)的一系列谱系,拥有11个染色体水平的全新组装的基因组和476个转录组样本。我们发现,木本竹类亚基因组表现出惊人的核型稳定性,在两个四倍体类群中表现出并行的亚基因组优势,在六倍体类群中逐渐转移优势。异源多倍化和亚基因组优势塑造了木本竹的木质竹秆的进化,以及快速生长和同步开花等特征。总的来说,该研究提供了对一个非凡的多倍体系统的基因组优势的见解,包括它对基因组环境的依赖以及它在进化时间中切换亚基因组优势的能力。
参考文献
Ma PF, Liu YL, Guo C, Jin G, Guo ZH, Mao L, Yang YZ, Niu LZ, Wang YJ, Clark LG, Kellogg EA, Xu ZC, Ye XY, Liu JX, Zhou MY, Luo Y, Yang Y, Soltis DE, Bennetzen JL, Soltis PS, Li DZ. Genome assemblies of 11 bamboo species highlight diversification induced by dynamic subgenome dominance. Nat Genet. 2024 Apr;56(4):710-720. doi: 10.1038/s41588-024-01683-0. Epub 2024 Mar 15. PMID: 38491323; PMCID: PMC11018529.
