文:回溯档案
编辑:回溯档案

植物中越来越多的新miRNA被发现,迄今为止,已在拟南芥、水稻和其他植物物种中注释了1000多个编码miRNA的基因,此外,还鉴定了其他几类小RNA,其特征在于其起源和生物学功能。
发育野生型和突变型胚珠

细胞分化伴随着转录、翻译和许多其他生理过程的变化,棉纤维是单细胞表皮毛状体,为研究拟南芥叶毛状体中也发生的细胞和发育事件提供了出色的模型。
纤维细胞的发育是一个复杂的形态和分子过程,其特征是细胞周期状态、转录控制和多种细胞骨架功能,包括集成的调控层次,最近,已经鉴定了许多控制早期纤维起始和晚期发育的基因,并且其中一些已经进行了功能表征。
GhMYB109是AtMYBGL1的推定直系同源物,在纤维细胞起始和伸长细胞中特异性表达,此外,GaMYB2的异位表达会在拟南芥种子的表皮中诱导单个毛状体,两个含有WD40结构域的棉花基因与拟南芥ttg1互补突变体。

对拟南芥叶毛和多基因表达的比较研究为棉花胚珠纤维发育提供了深入的见解,使用微阵列技术,对数百个转录本进行了分析,并在纤维细胞发育的早期阶段表现出不同的表达模式。
使用计算方法,我们最初从棉花中鉴定出37个潜在的miRNA;此外,还检测到了这些潜在miRNA的96个潜在靶点,最近,其他几个实验室也

这些研究报告的假定目标包括转录因子、生长素响应蛋白和其他与纤维发育相关的基因,为了探索小RNA在棉花纤维中的作用,最近分析了发育棉花胚珠的花后0-10天的小RNA序列,并获得了数百个小RNA。
尽管获得了583个小RNA的独特序列特征,但仅检测到了两个保守的miRNA,传统方法很可能测序深度不够,无法对胚珠中小RNA的全部复杂性进行采样,最近开发的高通量测序技术提供了一种强大的方法来识别和量化sRNA/miRNA。

小RNA最好通过具有高灵敏度和特异性的深度测序方法来发现和测量,此外,在基因组序列尚未完成的生物体中探索或注释miRNA是可行的,在这里,我们采用Solexa开发的深度测序方法来鉴定棉花胚珠中的小RNA,并分析小RNA的丰度和复杂性。
我们构建了两个分别由野生型和绒毛/无绒棉花胚珠制备的小RNA文库,样品是从0-10DPA发育的棉花胚珠中采集的,其中涵盖了主要的形态变化以及几个潜在的发育过程,包括纤维起始和伸长。

每个文库均单独测序,生成超过6-700万个短序列,总共超过1300万个序列读数,我们从棉花胚珠中获得了代表36个家族的100多个保守miRNA,其中许多是在这项研究中最初发现的此外,还鉴定出了两个非保守的新型miRNA候选者。
序列分析
棉纤维的发育是一个复杂的过程,涉及大量的基因表达和调控,为了了解小RNA是否参与该过程,我们使用了陆地棉品种徐州142和徐州142背景中的无绒无绒突变体,两者都经过基因鉴定,并且除了突变种子的特征是很少或没有纤维。

转录本的微阵列分析表明,野生型和fl突变体胚珠中的许多基因在0-10DPA之间差异表达,基于这一事实,我们推断,对卵泡小RNA进行深度测序将提供小RNA成分的完整视图以及野生型和突变体之间小RNA的差异表达谱。
因此,从野生型和突变型胚珠构建了两个小RNA文库,对文库进行深度测序,分别从野生型和突变型胚珠中生成7,055,692和6,517,694个序列读数。

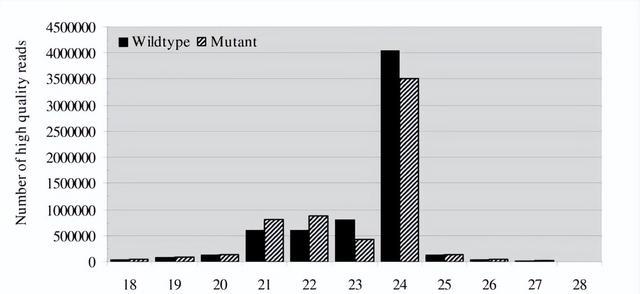
去除低质量读数和损坏的接头序列后,野生型保留6,584,945个读数,突变体保留6,069,470个读数,两个文库的大多数小RNA均为21-24nt,其中24nt小RNA最为丰富,这在Dicer衍生产品的典型尺寸范围内,并且与之前的大多数结果一致。
新miRNA候选物的分析
由于棉花基因组的完整测序无法获得,因此将独特的小RNA序列映射到棉花TIGR植物转录组合物序列,以鉴定潜在的新型miRNA,此外,由于棉花小RNA群体背景未知,秘密鉴定非保守miRNA相当具有挑战性。


miRNA靶标的预测
预测了所有已识别的miRNA家族的靶点,总共为21个保守miRNA家族预测了120个独特的靶基因,仅呈现具有已知或推定功能的基因,一些miRNA家族具有多个靶位点,表明这些miRNA在功能上存在差异。
此外,单个基因可能会被多个miRNA靶向,根据UniProt 描述的生物学功能,这些靶基因可分为10类,大多数靶标属于转录调控类别,表明这些基因编码转录因子。

其他几个组分别包含调节运输、氧化还原、信号转导途径和参与代谢的酶的基因,不幸的是,尚未预测这两种新候选miRNA的靶基因,这很可能归因于棉花基因组不完整。
迄今为止,已有一千多个植物miRNA基因被注释,其中一些已经得到了很好的表征,然而植物miRNA的数量似乎尚未饱和,植物物种中许多其他功能性miRNA仍有待研究。

与来自拟南芥和水稻的注释miRNA相比,来自棉花植物的miRNA很少被鉴定,最近进行了几项计算机模拟鉴定来自陆地棉的miRNA的研究,检测到大约18个高度保守的miRNA家族,并发现了几个不太保守的miRNA。
大部分可以通过深度测序恢复,只有一小部分不能通过深度测序恢复,这些缺失的miRNA可能归因于以下事实,本研究首次对陆地棉胚珠的小RNA群体进行深度测序,棉纤维细胞在胚珠中起始和发育,检测到数百万个长度为18~28nt的独特siRNA序列,对这些miRNA的进化保守性分析揭示了属于22个家族的111个保守miRNA。

连同miRBase中存在的几个陆地棉miRNA,本研究将把陆地棉中miRNA的数量提高到120,棉花胚珠中绝大多数保守的miRNA并不令人意外,本研究中鉴定的大多数miRNA在拟南芥中是保守的,只有少数在其他植物物种中是保守的,这种现象可以通过以下事实来解释。
棉纤维和表皮毛在表型上相似;两种类型的毛状体都使用共同的机制,例如,与调节毛状体起始和发育的转录因子密切相关。

值得注意的是,一些高度保守的miRNA家族,如miR156/157、miR167和miR172在单个文库中被测序超过一万甚至十万次,这些高度保守的miRNA可能代表了进化保守性和表达丰度之间的关系。
相反,一些不太保守甚至物种特异性的miRNA家族的丰度非常低,从进化的角度来看,这些miRNA在建立和维持不同生物群体之间的表型多样性方面发挥着作用,并参与谱系特异性途径和功能的调节。

除了保守的miRNA之外,本研究中鉴定的两个假定的miRNA在拟南芥和其他物种中没有直系同源物,由于非保守miRNA通常在特定细胞类型中以低水平表达。
表明这些棉花特异性miRNA可能在单子叶植物和双子叶植物谱系分化后扩展,支持了以下假设:每个物种的miRNA组都在快速且独立地进化。

突变体中miR398的表达远低于野生型,此前,拟南芥中的miR398被鉴定为编码Cu/Zn超氧化物歧化酶的靶基因,棉花胚珠中的miR398也被预测为类似的靶标。
有趣的是,最近在发育中的棉纤维的次生细胞壁中检测到了特定的棉花铜/锌超氧化物歧化酶,并表明其参与细胞壁生长吗,miR398是否调节超氧化物歧化酶和细胞壁生长将是一个值得研究的有趣课题。

结论
本研究首次对棉花纤维起始和发育的陆地棉胚珠的小RNA群体进行深度测序,检测到数百万个长度为18~28nt的独特miRNA序列,我们的结果支持了miRNA在调节不同细胞类型发育中的重要性,并表明在棉花纤维细胞中鉴定一套全面的miRNA将有助于我们了解纤维细胞起始和伸长的调节机制。
使用深度测序方法鉴定了棉花胚珠中的许多保守miRNA,我们的结果表明,野生型和突变型胚珠的miRNA存在差异表达谱,这有望调节明显参与棉纤维发育的转录本,进一步鉴定胚珠中这些差异表达的miRNA将有助于更好地了解纤维细胞发育的调控机制。
参考文献
【1】HeL,HannonGJ:小RNA在基因调控中发挥重要作用,纳特·吉内特(NatRevGenet),2004,5:522-531。
【2】Griffiths-JonesS、SainiHK、vanDongenS、EnrightAJ:miRBase:microRNA基因组学工具,核酸研究,2008,36(数据库问题):D154-D158。
【3】ChapmanEJ、CarringtonJC:内源性小RNA途径的专业化和进化,纳特·吉内特(NatRevGenet),2007,8:884-896。
【4】KimHJ,TripletBA:棉纤维在植物体内和体外的生长:植物细胞伸长和细胞壁生物发生的模型,植物生理学,2001年,127:1361-1366,10.1104/第010724页。
【5】王S,王JW,于N,李CH,罗B,苟JY,王LJ,陈XY:棉纤维MYB基因控制植物毛状体发育,植物细胞,2004,16:2323-2334。
