导语
鸟类性染色体在性别决定中发挥着独特的作用,影响鸟类的性形态和行为。核心水鸟是鸟类的一个主要类群,它们的共同特征是雌雄同构,性别间冲突水平较低,然而目前对于它们的性染色体演化方面仍然缺乏研究。朱鹮(Nipponia nippon)是一种典型的核心水鸟,属于鹈形目鹮科,体型较大,性单态,因此很难仅根据外表区分其性别,此外,朱鹮是一夫一妻制,没有明显或夸张的求偶行为,双亲共同抚育后代,包括筑巢、孵化、育雏。其他鸟类中,双亲投资的性别差异被认为在决定性选择的程度方面起着主要作用。朱鹮的性选择压力小,两性之间的性对抗可以忽略不计,为研究核心水鸟性染色体的进化提供了一个理想的模型。关于朱鹮独特的新性染色演化的相关研究发表在了《Nature Communications》期刊上。通过对朱鹮的雌性个体进行染色体水平组装的分析,确定了由微小染色体与古老性染色体融合而成的新型性染色体。解析了朱鹮性染色体的演化机制,为鸟类性染色体的演化历史和表达模式提供了新的见解。让我们一起来学习一下吧~

文献标题:Evolution and expression patterns of the neo-sex chromosomes of the crested ibis
发表期刊:Nature Communications
影响因子:16.6(Q1)
发表时间:2024.02.23
研究思路
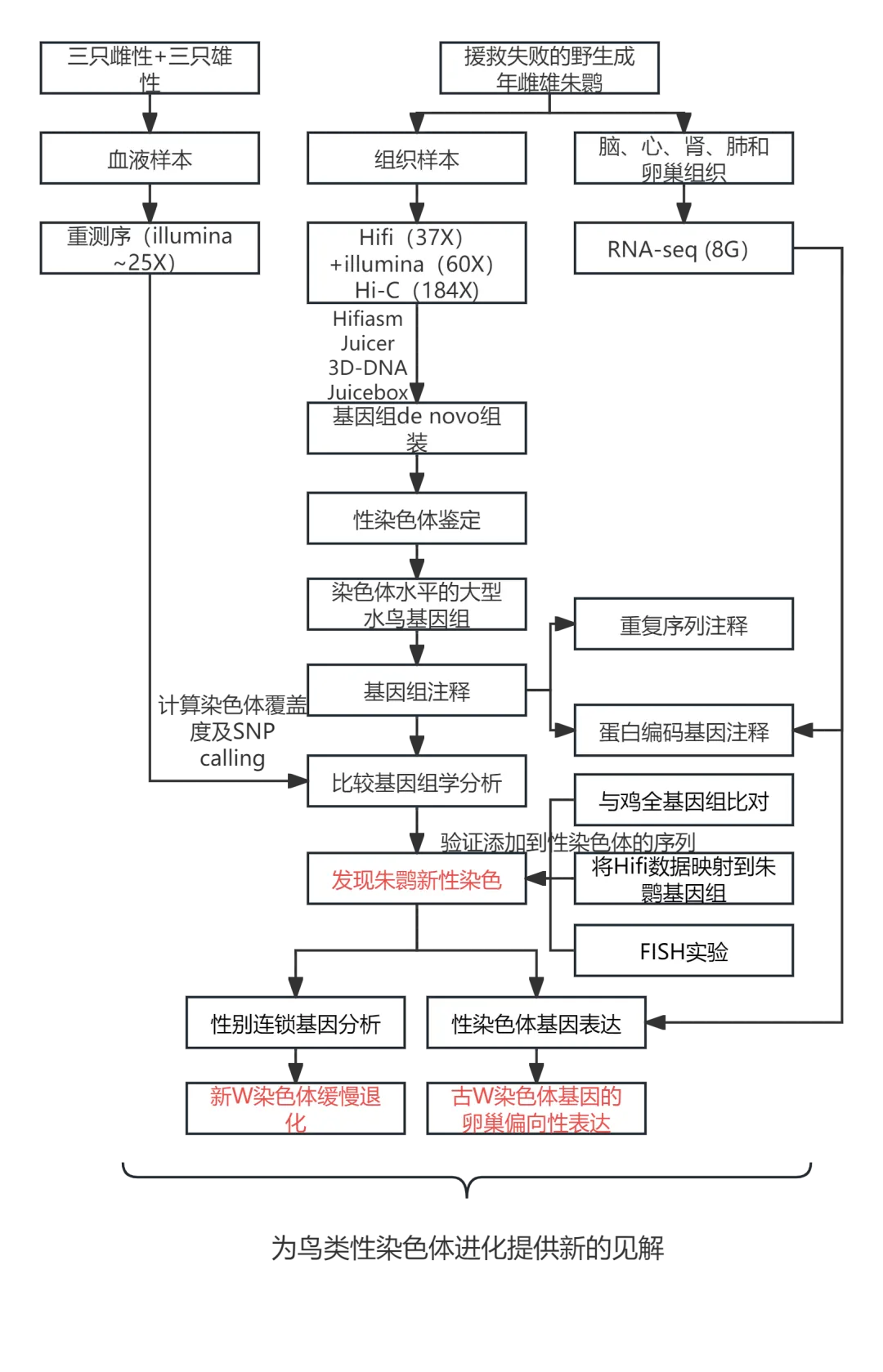
主要研究结果
1. 一个雌性朱鹮的新染色体水平基因组组装
利用PacBio HiFi reads(~ 37 x)、短读长reads (~ 60 x)和染色质构象捕获(Hi-C)读取(~ 184 x),重新组装了雌性朱鹮的高质量基因组,这是首次发表的核心水鸟染色体水平的基因组。N50全长16.4 Mb,比之前的基因组草图大630倍。该基因组有超过95.8%的contigs固定在29条染色体模型上(图1a),包括9条大染色体(chr1-9),18条微染色体(chr10-27)和一对ZW性染色体。根据已知的朱鹮核型(2n = 68),本次组装基因组仍缺少6条染色体模型。缺失的染色体可能是点状微染色体,可以通过超长ONT reads更好地分辨。该组装的基因组全长1.31 Gb,具有较高的连续性和完整性(BUSCO: 97.7%),与其他已发表的具有顶级组装质量的鸟类基因组相似。
通过比较雄性和雌性之间的测序覆盖率来确定Z和W染色体。在雌性中,Z和W染色体的测序覆盖率都是常染色体的一半,而在雄性中,W染色体的测序覆盖率很低(图1b)。组装的Z染色体是第六大染色体,包含1112个基因。与雌性相比,雄性Z染色体的测序覆盖率均匀地提高了2倍(图1b),这与对分化的Z染色体的预期一致。
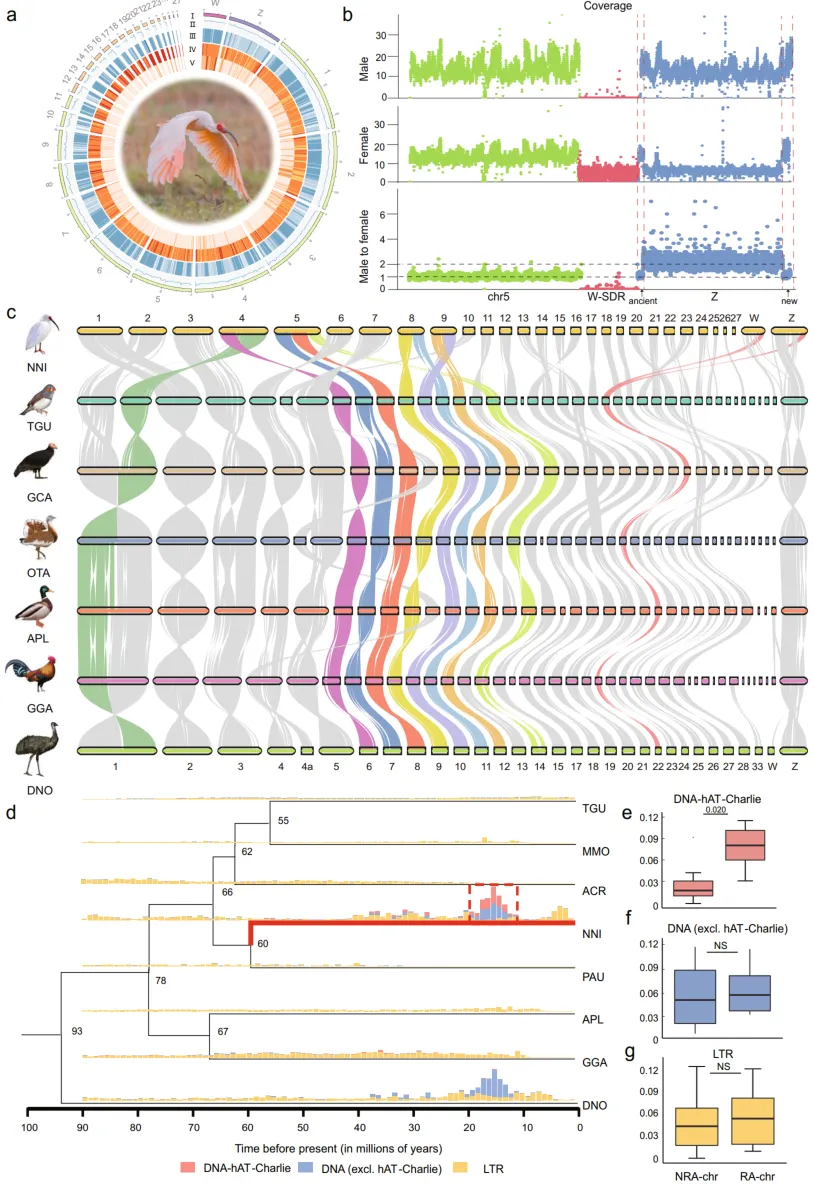
2. 频繁的染色体间重排可能与TE活性有关
为了研究朱鹮的染色体进化,对朱鹮(NNI)和几种代表性鸟类的基因组进行了多方位的同源性比较,包括栗耳草雀(Taeniopygia guttata, TGU)、加州兀鹫(Gymnogyps californianus, GCA)、大鸨(Otis tarda, OTA)、绿头鸭(Aanas platyrhynchos, APL)、鸡(Gallus gallus, GGA)和鸸鹋(Dromaius novaehollandiae, DNO)。分析显示,朱鹮基因组的染色体经历了至少六次融合事件,但没有发生分裂,这与其他具有大量染色体变化的鸟类相似(图1c)。在这些融合事件中,有四次是在大染色体和微染色体之间发生的,这在鸟类中是最常见的融合形式。而在鸟类中,大染色体之间的融合则较为罕见,但朱鹮中观察到了两次(在染色体1a和6之间(chr1a+chr6)以及染色体7和8之间(chr7+chr8))。
接下来,探究了是否存在某些转座子家族与朱鹮的染色体变化有关。结果显示,在朱鹮基因组中特定的DNA转座子家族(DNA-hAT-charlie)出现了物种特异性的大爆发,这些转座子在具有染色体间重排的染色体上显著富集(图1d, e)。相比之下,其他转座子插入不表现出对任何特定染色体的显著偏好(图1f, g,)。DNA-hAT-charlie元件大约在1500万年前开始在冠羽鹮的基因组中扩散(图1d)。
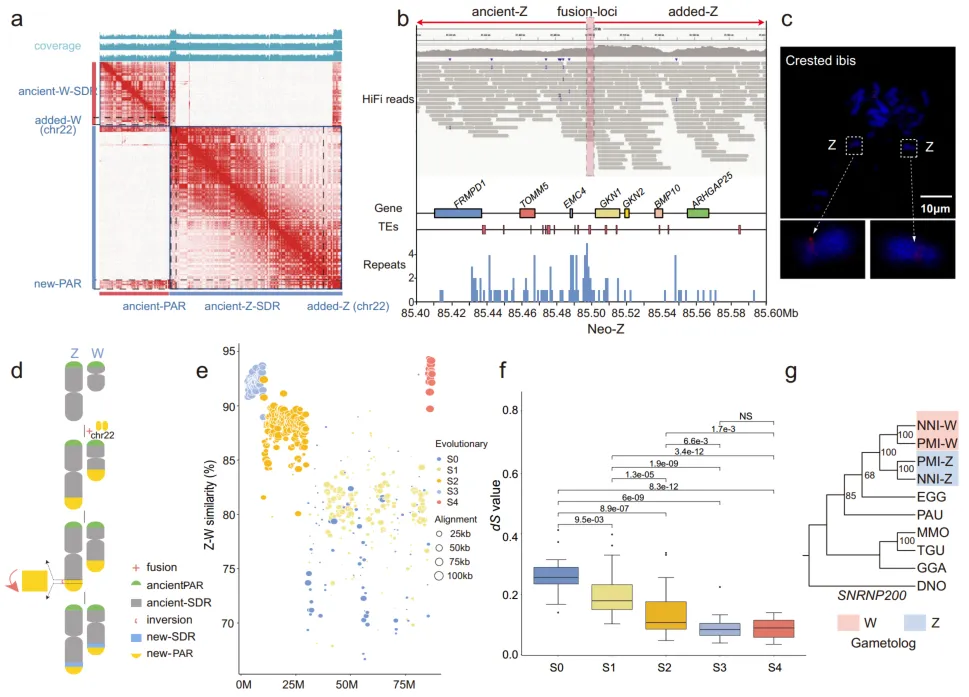
3. 朱鹮中新性染色体的形成
在六个染色体融合事件中,有一个似乎是染色体22(chr22)加入到古老的Z和W染色体上(图1c)。染色体22是一个微小染色体,其整个长度都添加到了古代性染色体上。这些是在核心水鸟类中发现的首个新性染色体。为了验证朱鹮的新Z/W染色体,仔细检查了古Z/W和新Z/W(染色体22来源部分)之间的融合点,分别位于新Z染色体的约85.5 Mb和新W染色体的约37.84 Mb处。Hi-C互作图强烈支持了组装的染色体模型(图2a)。此外,对于新Z和新W融合点,一个单一的contig跨越了相邻区域,并且这些contig被长测序读取均匀覆盖(图2b)。此外,使用特定的鸡染色体22序列探针进行的FISH实验显示,在一只朱鹮的Z染色体对上存在荧光信号,进一步验证了新性染色体是由古代性染色体和染色体22之间的融合事件导致的(图2c)。
4. 新性染色体上进化层的演变
性染色体间重组的独立抑制导致了三个进化阶段(S0–S2),这些阶段在所有新鸟小纲(Neoaves)鸟类中都是共享的。大多数Neoaves谱系都独立演化出一个额外的阶段S3,其大小各不相同。分析了新Z和W染色体之间的分化水平,确认了古代性染色体中的四个阶段(S0–S3),但也确定了新添加性染色体中的一个新阶段(S4)(图2e)。此外,S4在新W和新Z之间显示了约92.5%的序列相似性,类似于S3(91.9%),但高于较旧的阶段(S0和S1:80.7%;S2:87.5%,图2e)。S4的同义置换率(dS)与较旧的阶段相比较低,但与S3阶段相似(图2f),这表明S4的出现时间接近于S3。
为了调查新性染色体是否仅存在于冠羽鹮中,进行了鸡和另一种鹮类——铅色鹮(Theristicus caerulescens, TCA)之间的拟合度分析。在铅色鹮中发现了由chr22和古代-Z融合的新Z染色体,代表了鹮类中常见性染色体-常染色体融合事件。此外,对S4阶段的配子基因进行了系统发育分析,以及来自各种鸟类物种的其等位基因,包括铅色鹮和黑脸琵鹭(Platalea minor, PMI),后者属于Threskiornithidae的Platalea属。此分析结果显示,Threskiornithidae基因按染色体而不是按物种聚类,暗示了新添加的性染色体可能是在Threskiornithidae的共同祖先中形成的(图2g)。
研究总结
总的来说,该研究中对朱鹮的性染色体进行的分析揭示了与陆地鸟类先前发表的结果不同的性染色体进化特征,突显了鸟类性染色体多向演化的特点。该研究通过组装一个高质量的雌性朱鹮染色体水平的参考基因组,采用比较基因组学的方法,鉴定出朱鹮的22号常染色体和古性染色体融合形成的新性染色体,这是首次在核心水鸟中的报道。并且通过转录组学研究发现了朱鹮的新W染色体表现出缓慢降解的和明显的卵巢偏向性基因表达。这些发现为研究鸟类性染色体进化提供新的见解。
参考文献
Xu L, Ren Y, Wu J, Cui T, Dong R, Huang C, Feng Z, Zhang T, Yang P, Yuan J, Xu X, Liu J, Wang J, Chen W, Mi D, Irwin DM, Yan Y, Xu L, Yu X, Li G. Evolution and expression patterns of the neo-sex chromosomes of the crested ibis. Nat Commun. 2024 Feb 23;15(1):1670. doi: 10.1038/s41467-024-46052-x. PMID: 38395916; PMCID: PMC10891136.
